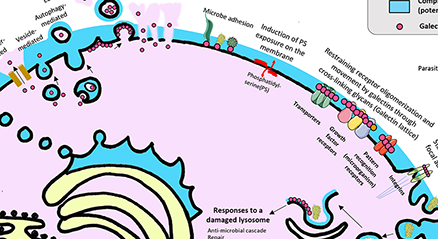
ガレクチンは、糖鎖認識ドメイン(Carbohydrate Recognition Domain: CRD)に特徴的なアミノ酸配列をもつ糖結合タンパク質である。ガレクチンファミリーの多くのメンバーは、その他に以下の2つの特徴も示す。1つは、細胞の内側にも外側にも存在すること、もう1つは、タンパク質間相互作用を介して複数のパートナーと結合することである。細胞内では、様々なガレクチンの非糖鎖リガンドの例として以下が挙げられる。(a)プロト型のガレクチン-1は、がん遺伝子のH-Rasや転写因子のOCA-B及びTFII-Iに結合する。(b)タンデムリピート型のガレクチン-8は、ガレクチン-9などの他のガレクチンや、オートファジー受容体NDP52、TRIM5α(Tripartite Motif 5α)に結合する。(c)キメラ型のガレクチン-3のNH2末端ドメインは、Tsg101等の輸送に関与するエンドソーム輸送選別複合体(endosomal sorting complex required for transport、ESCRT複合体)の因子やhnRNP A2B1などのリボ核タンパク質複合体に結合する一方、COOH末端ドメインはアポトーシス抑制因子Bcl-2、TRIM16、そして転写因子OCA-B、TFII-I及びβ-カテニンに結合する。本稿では、細胞内ガレクチンの浮気性(promiscuity)を浮き彫りにした研究の一部と、これらの発見により生じたいくつかの興味深い疑問を要約する。 ...and more
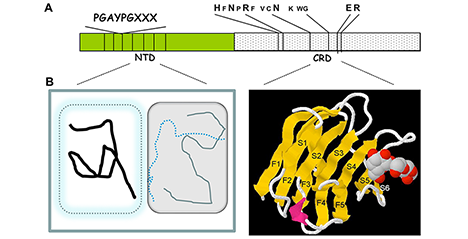
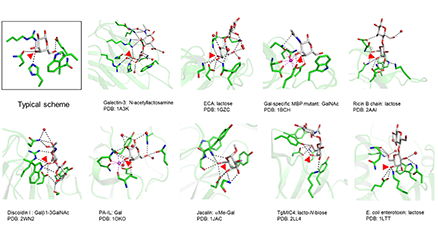
ガレクチンの天然リガンドは、主に糖タンパク質であり、スフィンゴ糖脂質ともおそらくある程度結合する。遊離状態の(編集者注:タンパク質や脂質を付加されていない)オリゴ糖に対するガレクチンの結合親和性および特異性は広く研究されているが、本来のリガンドである糖タンパク質や糖脂質に付加した状態の糖との結合についてはあまり調べられていない。糖鎖に対するガレクチンの結合性は、遊離オリゴ糖として検討した場合よりも、糖タンパク質や細胞表面に存在する状態で検討した場合のほうが、見かけの親和性が高くなることが多い。本稿では、このような現象を生じる主な要因が多価相互作用ではなく、ガレクチン-糖認識ドメイン(carbohydrate recognition domain:CRD)が提供する広範囲にわたる結合部位の寄与により一価性の親和性がより高くなることが重要であることを、その証拠を提示しながら論じる。また、このような一価の相互作用は、高い選択性を有している場合もあり、この選択性は結合する糖鎖の構造および糖鎖付加部位付近のタンパク質部分の両方によってもたらされる。これに対し、多価相互作用はさまざまな形でリガンドを架橋形成することができるのでガレクチンの機能を発揮する上で不可欠である。一価相互作用によって生じる選択性に依存している架橋形成であるが、架橋によりさらにもう1段階の選択性や親和性の追加に寄与する場合もある。本稿では主にガレクチン-1および-3に焦点を当てるが、他のガレクチンについても言及する。細胞内におけるガレクチンと糖鎖が付加されていないタンパク質リガンドの相互作用も同様に興味深いが、ここでは取り上げない。 ...and more

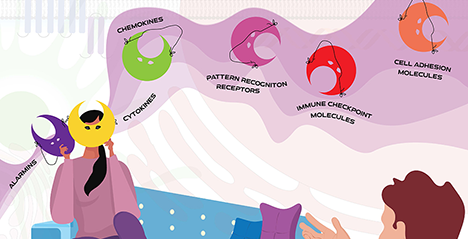
多重人格障害は、異なる自己人格が自発的に発生されるという特徴を持つ精神疾患である。各人格はそれぞれに特有の形で現れ、別々の名前や生活圏、ライフスタイルを有していることが多い。ある人格から別の人格への交代は、一般的に環境および対人的要因によって引き起こされる1。この疾患の特徴は、まさにガレクチンの持つ性質に似ている。進化的に保存された糖結合タンパク質ファミリーであるガレクチンは、特定の機能に特化している多くの細胞内分子および経路とは異なり、免疫において多様な機能・役割を持っているからである。ガレクチンファミリーに属するタンパク質は、その多くが細胞内の異なるコンパートメント(核、細胞質、およびオルガネラ)間を行き来し、細胞外環境に放出される。そして、低酸素、栄養状態、細胞内外のpH、サイトカイン環境、炎症性または免疫抑制性シグナルの存在など、多様な微小環境刺激に応答して異なる役割を獲得する。免疫系におけるガレクチンは、自然免疫および獲得免疫応答を調整する重要な機能を幅広く発揮することができ、正常および疾患状態における免疫細胞の協調的な振る舞いを作り出す上で重要な役割を担っている。興味深いことに、同じガレクチンであっても、活性化、分化、免疫細胞遊走などの細胞プログラムの違いや、病原体の侵入、自己免疫による炎症、線維症、がんなどの病態によって、サイトカイン、ケモカイン、細胞接着分子、免疫チェックポイント分子、危険・ダメージ関連分子パターン(danger-associated molecular pattern: DAMP)、あるいは成長因子として機能しうる。 ...and more
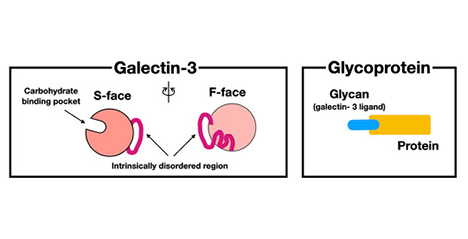
ガレクチン-3はガレクチン家系において、ガレクチン-1と共に最も研究されているガレクチンである。ガレクチン-3がin vitroでは、細胞内および細胞外で、そしてin vivoにおいてはさまざまな器官の機能に関与していることが報告されており、その範囲は多岐に及ぶ。前編で述べた様に、ガレクチン-3はそのN末端に、他の動物レクチンには見られないペプチドの繰り返し構造が続く「天然変性領域」を含む領域を含み、そのペプチド長はC末端のレクチンドメイン(Carbohydrate-binding domain: CBD)とほぼ同じである(図 3)。報告されているガレクチン-3の活性の多くはこの天然変性領域に依存している。また、通常は単量体で存在するガレクチン-3は糖鎖リガンドと結合したのちに、天然変性領域に依存して多量体化する。従って、ガレクチン-3の活性のほとんどは、天然変性領域が仲介する多量体形成によって調節されていると考えられている。では、他のレクチンに見られないこのユニークな天然変性領域がどのようにガレクチン-3の多量体化に関与しているかについて、後編では最近の知見を取り入れながら、さらに掘り下げ、そこから多量体化とガレクチン-3の多岐にわたる機能との関係について探っていきたい。 ...and more
ガレクチン-3は他のガレクチン家系メンバーと構造的に全く異なる。C末端のβ-ガラクトシドに親和性のあるレクチンドメイン(Carbohydrate-binding domain: CBD)に加えN末端にCBDとほぼ同じ長さの非レクチン部分がガレクチン-3には存在するからだ。非レクチン部分の大部分はプロリン、グリシン、チロシンに富んだ9〜13個のアミノ酸からなるペプチドが9〜12回繰り返した構造で、特定の二次および三次構造を形成しない「天然変性領域」である.。過去数年の研究で、糖鎖に結合したガレチン-3がこのN末の変性領域を介して液–液相分離(Liquid-liquid phase separation、以下LLPS)に関与している可能性が示されてきた。 ...and more
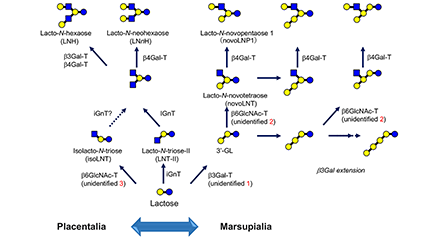
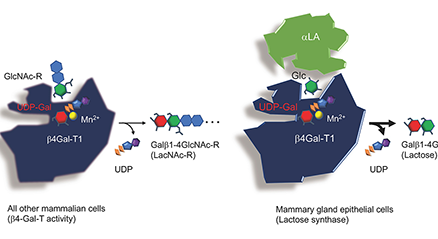
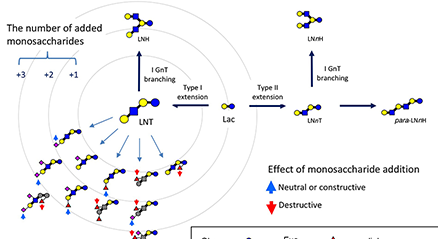
ヒトの乳に12~13 g/Lの濃度で含まれるオリゴ糖(ヒトミルクオリゴ糖, HMOs)は、これまでに約250種類が分離され、160種類以上の化学構造が決定されている。それらはフコースやシアル酸を含まないコア骨格構造に基づいて、20のシリーズに分類されるが、コア骨格の生合成系はiGnT, IGnT, β3GalT, β4GalTの活性に基づいて提案されている。また、ウシの初乳からも約50種類のミルクオリゴ糖が分離・構造決定されているが、それらのコア骨格の生合成経路も提案されている。 ...and more
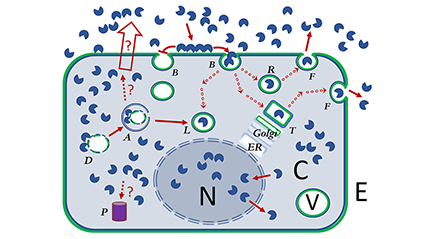
ガレクチンは小さな可溶性の糖鎖結合タンパク質からなる家系であり、数多くの細胞機能に関与することが知られている。具体的には、炎症や線維症、がんなどの疾患に対する診断や治療標的として注目を集めている。しかしながら、ガレクチンの役割は枚挙に暇がなく、ありとあらゆる生命現象(コンテクスト)に絡んでいるのが事実である。例えば、ガレクチンは細胞内でも細胞外でも、低濃度でも高濃度でも作用し、また、緩徐にも迅速にも機能し、低い特異性で働くこともあれば高い特異性での機能もある。このため、「これがガレクチン家系の、あるいは特定のガレクチンの機能である」という具合にガレクチンの機能を一言で言い表すのは難しい。本稿では、ガレクチンに残された、興味深く、かつ重要な未解決問題を読者に提起するために、上記の現象を具体的に述べながら、ガレクチンの一生を論じてみたい。ただし、ここで述べる事柄は必ずしも正確を期したものではない。とは言え、内容は概ね正しく、ガレクチンにまつわる多くの疑問を明示するのに役立つだろう。...and more
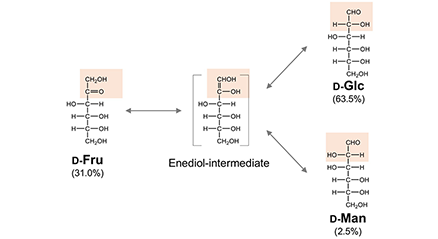
第1部ではガレクチンが細胞質タンパク質である点に着目し、その起源について述べた。本章では、「糖が存在しなければレクチンは存在しない」との仮定の下で議論を進める。この前提に従えば、ガレクチンの誕生には、それに先行する糖リガンド、すなわちガラクトースの存在が不可欠となる。では、ガラクトースはいつ、いかにして発生したのか。本論では、著者が提唱した糖の起源仮説における「ガラクトース後生説」をベースに、その概略を述べる(詳細は文献1-4を参照)。...and more
ガレクチンの特性を端的に言いあらわせば進化的に関連したガラクトース(厳密にはβガラクトシド)結合性タンパク質である。この分子群の生物分布や多様な機能については各所で述べられているが、その起源についてはほとんど議論されたことがない。ガレクチンは糖に結合するという固有の性質上、細胞外での機能が想定されるが、今まで調べられたガレクチンすべてが分泌のためのシグナル配列を持たない。実際、ガレクチンはどの生物由来であっても「細胞質タンパク質」として生合成される。このことは、ガレクチンが本来細胞質で何らかのミッションを担っていた可能性を示唆する。本論(第一部)では、ガレクチンの起源を細胞質タンパク質に求め、これについて論ずる。さらに、ガレクチンの機能を論ずる際不可欠となるガラクトースの「そもそも論」を取り上げる(第二部)。すなわち、ガラクトースの起源について議論するとともに、糖やキラリティーの起源について言及する。...and more
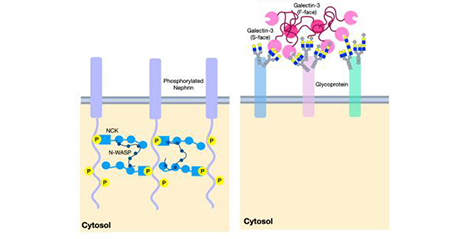
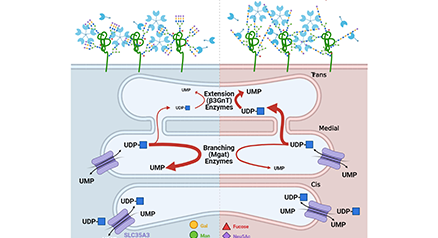
ガレクチン格子は、ガレクチンと細胞表面に存在する糖タンパク質間の多価の結合によって生じる構造体である。 結合パートナーをすばやく入れ替えることができ、このため、膜上を自由に拡散する糖タンパク質と、安定した糖タンパク質複合体との中間的構造―すなわち液-液相分離としての特徴を併せもつ。ガレクチン格子は、(i)受容体や溶質トランスポーターの被覆ピットを介したエンドサイトーシス(coated-pit endocytosis)やカベオリン領域への流れを制御し、(ii)免疫シナプスや焦点接着複合体(focal adhesion complexes)のような細胞間接触の機会創出を演出している。ゴルジ体におけるN型糖鎖の構造変化(リモデリング)を起こすようなUDP-GlcNAc供給に変化が起こると、ガレクチンの糖タンパク質への親和性にも変化が起こり、ひいてはガレクチン格子による細胞表面の糖タンパク質の動態にも変化が生じる。さらにUDP-GlcNAc供給は代謝制御を受けている。この格子モデルは、マウスの免疫制御、がんの進行、グルコースホメオスタシスといった現象において立証されている。本稿では、代謝、ガレクチン、および糖タンパク質リガンドの間に存在する相互作用に加え、炎症や自己免疫の予測・治療におけるこのモデルの有用性について概説する。...and more
通常、レクチンと糖の親和性はµMオーダーであるが、ミルク中には、オリゴ糖が10 mM以上、ラクトースにいたっては100 mMを超える濃度で糖が豊富に含まれる。この濃度は、レクチンの解離定数の観点からみると衝撃的な濃さであり、ガレクチンを含む多くのレクチンの糖結合活性は阻害されてしまうだろう。この阻害活性の生物学的な意味は何であろう?また、消化管内や体内でのミルクオリゴ糖濃度はどうなっているのだろう?と、いくつもの疑問がわいてくる。
そこで、今回は、ミルクオリゴ糖が生体内でどう利用され、どのような役割をはたすか、また、消化管に豊富に発現するガレクチンの機能との関係について、推測、妄想、議論を深めたい。そして、哺乳類がラクトース生合成能を獲得し、乳児の栄養源として利用したことが『哺乳類の成功の鍵』といわれる理由について、改めて考察してみたい。...and more
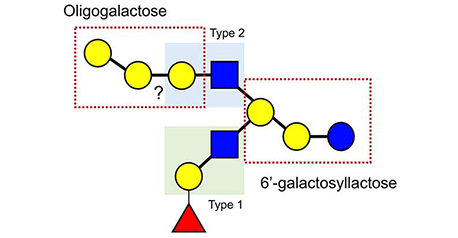
本編「ミルクオリゴ糖とガレクチン」(Glycoforum. 2021 Vol.24 (2), A3)の中で、ラットミルク中のオリゴ糖のほとんどは3’-SLと6’-SLであるのに対し、齧歯目のローランドパカ(Cuniculus paca)のミルクはそれ以外にもより複雑なミルクオリゴ糖を含んでいることを紹介した。このことから直観されるのは、実験動物のミルクオリゴ糖は多様性を失い、単純化しているのでは、という考えである。一方、最近極微量ではあるが、ラットやマウスの乳にも硫酸化した3’-SLなど他のオリゴ糖も発見されている。ミルクオリゴ糖の構造多様性はどのようにして決定しているのであろうか。「番外編」第2弾はこの点に絞って議論してみたい。...and more
本稿はグライコフォーラムVol. 24 (2)に 掲載の「ミルクオリゴ糖とガレクチン」(浦島、平林;以下、本編という)のspin-off、いわば「番外編」である。本編では賄いきれなかったテーマの延長上にある、さらなる議論や未解決課題について、ざっくばらんに、かつ自由、闊達な立場で著者らの仮説(妄想)を展開しようと思う。フォーラムという建付けをご理解の上、お付き合いいただきたい。 ...and more
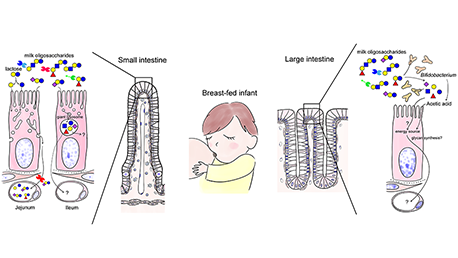
哺乳動物の乳/初乳は通常主要な糖質としてラクトース(Galβ1-4Glc)以外に、各種ミルクオリゴ糖を含んでいる。その大半はラクトース骨格を還元末端側に含み、多くはガラクトース(Gal)、N-アセチルグルコサミン(GlcNAc)、フコース(Fuc)、N-アセチルまたN-グリコリルノイラミン酸(Neu5AcまたNeu5Gc)、またN-アセチルガラクトサミン(GalNAc)を構造内に含んでいる。例えばヒト乳には、12〜13 g/Lの濃度注1)の、250種類以上のミルクオリゴ糖が含まれ、そのうち現在までに170種類近くの化学構造が決定されている1。本稿ではミルクオリゴ糖の概要紹介とともに、ガレクチンにまつわる未解決問題を解決するための独自の探索ツールとしてのミルクオリゴ糖の利用可能性を展望する。...and more
レクチンは、糖鎖にコードされた情報を生理活性に変換するプロセスにおいて中心的な役割を担っている。様々なレクチンは糖鎖認識ドメイン(CRD)の折り畳み構造に基づいて分類されるが、個々の構造単位は、一連の糖鎖と相互作用する以外にも多くの機能を有しているようだ。特定のアミノ酸配列エレメントはCRDの機能プロファイルを拡張することができるようであり、このことはガレクチンのCRDにおいて原理証明されている。つまり、特定の配列が、活性や立体構造、四次構造に対する分子スイッチとして機能するだけでなく、さらに、タイプの異なる結合パートナーに対して相補的領域を提供することも可能になる。その結果、CRDは思いも寄らない多機能性を獲得できるのだ。このような可能性をさらに検討・検証することで、「なぜガレクチンが、シグナル配列を持たずに分泌されるタンパク質の小さなグループに属しているのか」といった、重要な未解決問題の答えが得られるかもしれない。CRDが糖との結合に必要な部分以外にも、多様な構造エレメントで構成されている可能性を教えてくれる貴重な事例研究である。また、公衆衛生上重要な知見として、 コロナウイルス(SARS-CoV-2を含む)の宿主接着分子(ウイルス―スパイクタンパク質)にガレクチン様の折り畳み構造が含まれることが挙げられる。宿主遺伝子の一部をウイルスが取り込んだとすれば、上記議論は、我々を襲うパンデミックの脅威に対抗する革新的な治療法を見出す契機となるかもしれない。...and more
ガレクチンはずいぶん有名になったが、あいかわらずわかりにくい分子である。登場する舞台は多岐にわたるが、主役らしいふるまいはあまり見せない。天職は脇役のようだが、いないと絶対に困るメンバーだ。いろいろな相手とつきあうが、その条件は、素性(たんぱく質か、脂質か、プロテオグリカンか)や地位はどうでもよく、お気に入りの衣装(糖鎖)さえまとっていればよい。だから細胞外たんぱく質だったら、ほとんど(1万種類以上)が相手になりうる。ただし、あまり深いつき合い方はしない。ガレクチンの本質がとらえにくい原因のひとつが、こうした広くて浅い社交性にある。この講話では、こうした点を踏まえて、いくつか基礎的なことを再点検してみよう。 ...and more

このノートは、大腸菌を利用したヒトガレクチンファミリーの発現と精製に関する未発表データなどをまとめたものです。これまでガレクチンに馴染みがなく、新たに組換えガレクチンタンパク質の発現・精製を行う研究者(特に、生化学研究の経験が比較的浅い研究者)の参考になればと思います。 ...and more
この度、グライコフォーラムの新シリーズとして「ガレクチン」を開始する運びとなった。これは、昨今話題の多いガレクチンについて、実験プロトコールを関係者と広く共有するとともに、通常学術誌ではテーマとして取り上げることが難しい「未解決問題」を扱うという全く新しいコンセプトに基づく企画である。産総研の平林とラヴァル大学の佐藤が編集を務めさせていただく。 ...and more