Diehl C, Engstrom O, Delaine T, Hakansson M, Genheden S, Modig K, Leffler H, Ryde U, Nilsson UJ, Akke M. Protein flexibility and conformational entropy in ligand design targeting the carbohydrate recognition domain of galectin-3. J Am Chem Soc. 2010;132(41):14577-89.
Fenley AT, Muddana HS, Gilson MK. Entropy-enthalpy transduction caused by conformational shifts can obscure the forces driving protein-ligand binding. Proc Natl Acad Sci U S A. 2012;109(49):20006-11.
Verteramo ML, Stenstrom O, Ignjatovic MM, Caldararu O, Olsson MA, Manzoni F, Leffler H, Oksanen E, Logan DT, Nilsson UJ, Ryde U, Akke M. Interplay between Conformational Entropy and Solvation Entropy in Protein-Ligand Binding. J Am Chem Soc. 2019;141(5):2012-26.
Jencks WP. On the attribution and additivity of binding energies. Proc Natl Acad Sci U S A. 1981;78(7):4046-50.
Murray CW, Verdonk ML. The consequences of translational and rotational entropy lost by small molecules on binding to proteins. J Comput Aided Mol Des. 2002;16(10):741-53.
Carlsson S, Oberg CT, Carlsson MC, Sundin A, Nilsson UJ, Smith D, Cummings RD, Almkvist J, Karlsson A, Leffler H. Affinity of galectin-8 and its carbohydrate recognition domains for ligands in solution and at the cell surface. Glycobiology. 2007;17(6):663-76.
Zetterberg FR, Peterson K, Johnsson RE, Brimert T, Hakansson M, Logan DT, Leffler H, Nilsson UJ. Monosaccharide Derivatives with Low-Nanomolar Lectin Affinity and High Selectivity Based on Combined Fluorine-Amide, Phenyl-Arginine, Sulfur-pi, and Halogen Bond Interactions. ChemMedChem. 2018;13(2):133-7.
Lee YC, Townsend RR, Hardy MR, Lonngren J, Arnarp J, Haraldsson M, Lonn H. Binding of synthetic oligosaccharides to the hepatic Gal/GalNAc lectin. Dependence on fine structural features. J Biol Chem. 1983;258(1):199-202.
Lee RT, Lee YC. Affinity enhancement by multivalent lectin-carbohydrate interaction. Glycoconj J. 2000;17(7-9):543-51.
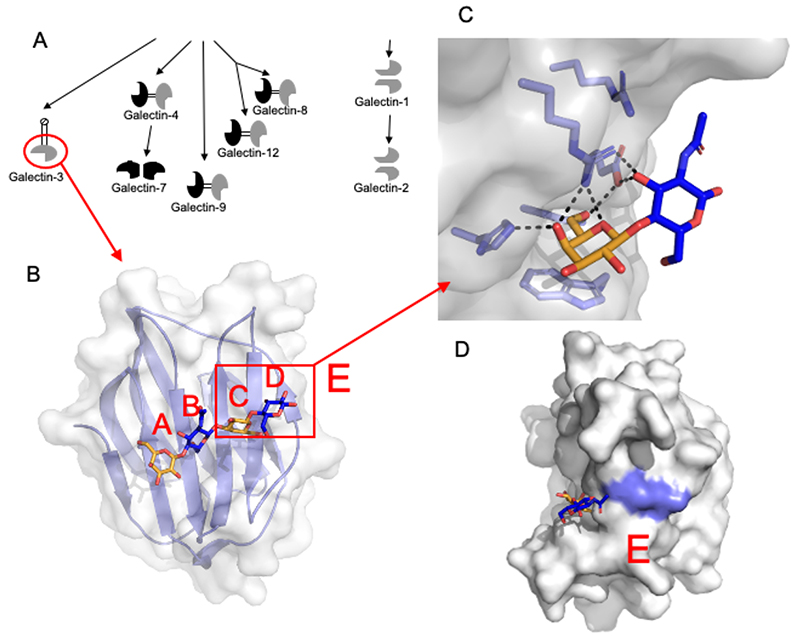
Leffler H, Carlsson S, Hedlund M, Qian Y, Poirier F. Introduction to galectins. Glycoconj J. 2002;19(7-9):433-40.
Leffler H, Barondes SH. Specificity of binding of three soluble rat lung lectins to substituted and unsubstituted mammalian beta-galactosides. J Biol Chem. 1986;261(22):10119-26.
Sparrow CP, Leffler H, Barondes SH. Multiple soluble beta-galactoside-binding lectins from human lung. J Biol Chem. 1987;262(15):7383-90.
Hirabayashi J, Hashidate T, Arata Y, Nishi N, Nakamura T, Hirashima M, Urashima T, Oka T, Futai M, Muller WE, Yagi F, Kasai K. Oligosaccharide specificity of galectins: a search by frontal affinity chromatography. Biochim Biophys Acta. 2002;1572(2- 3):232-54.
Stowell SR, Arthur CM, Mehta P, Slanina KA, Blixt O, Leffler H, Smith DF, Cummings RD. Galectin-1, -2, and -3 exhibit differential recognition of sialylated glycans and blood group antigens. J Biol Chem. 2008;283(15):10109-23.
Salomonsson E, Carlsson MC, Osla V, Hendus-Altenburger R, Kahl-Knutson B, Oberg CT, Sundin A, Nilsson R, Nordberg-Karlsson E, Nilsson UJ, Karlsson A, Rini JM, Leffler H. Mutational tuning of galectin-3 specificity and biological function. J Biol Chem. 2010;285(45):35079-91.
Iwaki JH, J. Carbohydrate-Binding Specificity of Human Galectins: An Overview by Frontal Affinity Chromatography. Trends in Glycoscience and Glycotechnology. 2018;30(172):SE137-SE53.
Johannes L, Jacob R, Leffler H. Galectins at a glance. J Cell Sci. 2018;131(9).
Johannes L, Shafaq-Zadah M, Dransart E, Wunder C, Leffler H. Endocytic Roles of Glycans on Proteins and Lipids. Cold Spring Harb Perspect Biol 2023;doi: 10.1101/cshperspect.a041398.
Finne J, Breimer ME, Hansson GC, Karlsson KA, Leffler H, Vliegenthart JF, van Halbeek H. Novel polyfucosylated N-linked glycopeptides with blood group A, H, X, and Y determinants from human small intestinal epithelial cells. J Biol Chem. 1989;264(10):5720- 35.
Iwaki J, Tateno H, Nishi N, Minamisawa T, Nakamura-Tsuruta S, Itakura Y, Kominami J, Urashima T, Nakamura T, Hirabayashi J. The Galbeta-(syn)-gauche configuration is required for galectin-recognition disaccharides. Biochim Biophys Acta. 2011;1810(7):643- 51.
Gao C, Hanes MS, Byrd-Leotis LA, Wei M, Jia N, Kardish RJ, McKitrick TR, Steinhauer DA, Cummings RD. Unique Binding Specificities of Proteins toward Isomeric Asparagine-Linked Glycans. Cell Chem Biol. 2019;26(4):535-47 e4.
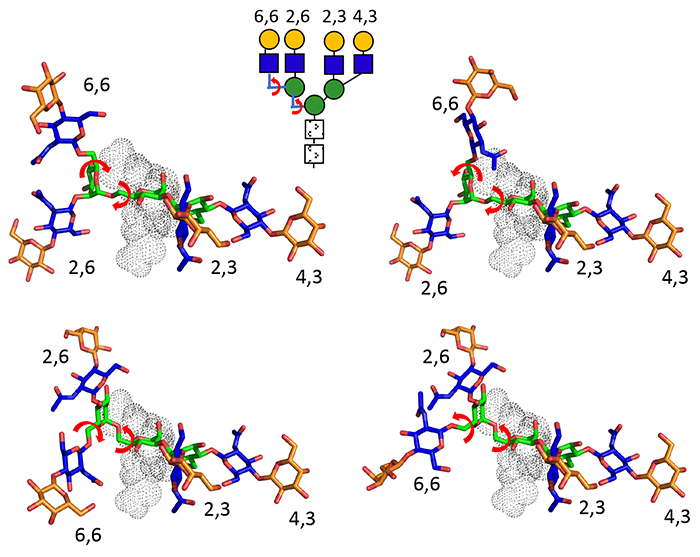
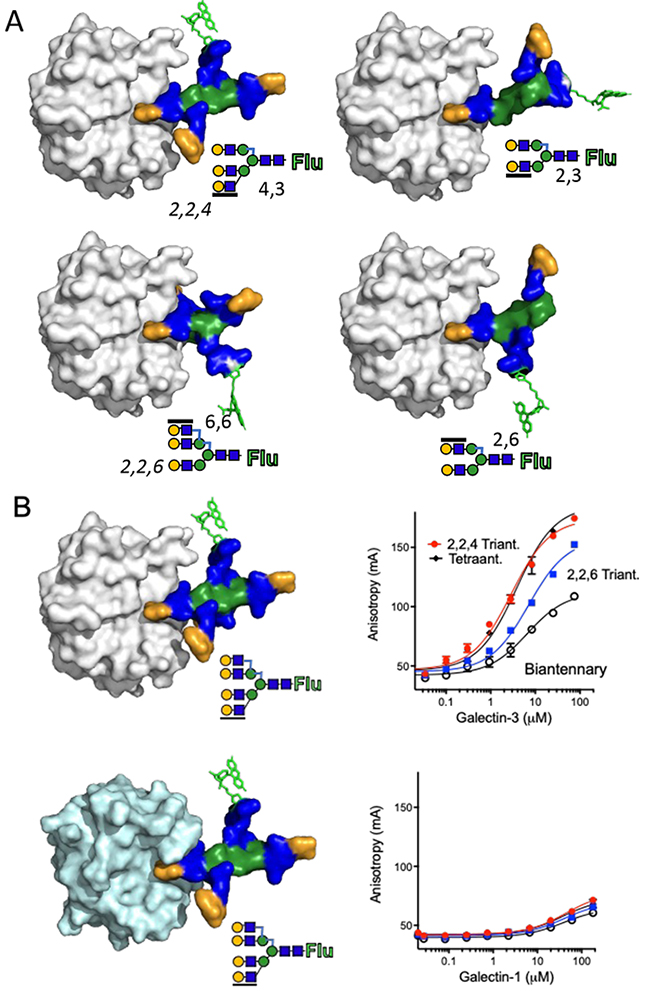
Patnaik SK, Potvin B, Carlsson S, Sturm D, Leffler H, Stanley P. Complex N- glycans are the major ligands for galectin-1, -3, and -8 on Chinese hamster ovary cells. Glycobiology. 2006;16(4):305-17.
Nielsen MI, Stegmayr J, Grant OC, Yang Z, Nilsson UJ, Boos I, Carlsson MC, Woods RJ, Unverzagt C, Leffler H, Wandall HH. Galectin binding to cells and glycoproteins with genetically modified glycosylation reveals galectin-glycan specificities in a natural context. J Biol Chem. 2018;293(52):20249-62.
Cederfur C, Salomonsson E, Nilsson J, Halim A, Oberg CT, Larson G, Nilsson UJ, Leffler H. Different affinity of galectins for human serum glycoproteins: galectin-3 binds many protease inhibitors and acute phase proteins. Glycobiology. 2008;18(5):384-94.
Carlsson MC, Bengtson P, Cucak H, Leffler H. Galectin-3 guides intracellular trafficking of some human serotransferrin glycoforms. J Biol Chem. 2013;288(39):28398-408.
Grothaus IL, Bussi G, Colombi Ciacchi L. Exploration, Representation, and Rationalization of the Conformational Phase Space of N-Glycans. J Chem Inf Model. 2022;62(20):4992-5008.
Dialpuri JS, Bagdonas H, Atanasova M, Schofield LC, Hekkelman ML, Joosten RP, Agirre J. Analysis and validation of overall N-glycan conformation in Privateer. Acta Crystallogr D Struct Biol. 2023;79(Pt 6):462-72.
Woods RJ. Predicting the Structures of Glycans, Glycoproteins, and Their Complexes. Chem Rev. 2018;118(17):8005-24.
Sato S, Hughes RC. Binding specificity of a baby hamster kidney lectin for H type I and II chains, polylactosamine glycans, and appropriately glycosylated forms of laminin and fibronectin. J Biol Chem. 1992;267(10):6983-90.
Carlsson MC, Cederfur C, Schaar V, Balog CI, Lepur A, Touret F, Salomonsson E, Deelder AM, Ferno M, Olsson H, Wuhrer M, Leffler H. Galectin-1-binding glycoforms of haptoglobin with altered intracellular trafficking, and increase in metastatic breast cancer patients. PLoS One. 2011;6(10):e26560.
Cunjie Zhang MS-Z, Judy Pawling, Deanna Wan Jie Ng, Geoffrey G. Hesketh, Estelle Dransart, Karina Pacholczyk, Joseph Longo, Anne-Claude Gingras, Linda Z. Penn, Ludger Johannes, James W. Dennis. SLC3A2 N-glycosylation and alternate evolutionary trajectories for amino acid metabolism. bioRxiv 2022;11.15.516651: https://doi.org/10.1101/2022.11.15.516651
Paz I, Sachse M, Dupont N, Mounier J, Cederfur C, Enninga J, Leffler H, Poirier F, Prevost MC, Lafont F, Sansonetti P. Galectin-3, a marker for vacuole lysis by invasive pathogens. Cell Microbiol. 2010;12(4):530-44.
Varki A, Cummings RD, Esko JD, Stanley P, Hart GW, Aebi M, Mohnen D, Kinoshita T, Packer NH, Prestegard JH, Schnaar RL, Seeberger PH. Essentials of glycobiology. Fourth edition. ed. Cold Spring Harbor: Cold Spring Harbor Laboratory Press; 2022. pages cm p.
Barboni EA, Bawumia S, Hughes RC. Kinetic measurements of binding of galectin 3 to a laminin substratum. Glycoconj J. 1999;16(7):365-73.
Leppanen A, Stowell S, Blixt O, Cummings RD. Dimeric galectin-1 binds with high affinity to alpha2,3-sialylated and non-sialylated terminal N-acetyllactosamine units on surface-bound extended glycans. J Biol Chem. 2005;280(7):5549-62.
Dimitroff CJ. I-branched carbohydrates as emerging effectors of malignant progression. Proc Natl Acad Sci U S A. 2019;116(28):13729-37.
Bian CF, Zhang Y, Sun H, Li DF, Wang DC. Structural basis for distinct binding properties of the human galectins to Thomsen-Friedenreich antigen. PLoS One. 2011;6(9):e25007.
Rodriguez MC, Yegorova S, Pitteloud JP, Chavaroche AE, Andre S, Arda A, Minond D, Jimenez-Barbero J, Gabius HJ, Cudic M. Thermodynamic Switch in Binding of Adhesion/Growth Regulatory Human Galectin-3 to Tumor-Associated TF Antigen (CD176) and MUC1 Glycopeptides. Biochemistry. 2015;54(29):4462-74.
Ideo H, Seko A, Ishizuka I, Yamashita K. The N-terminal carbohydrate recognition domain of galectin-8 recognizes specific glycosphingolipids with high affinity. Glycobiology. 2003;13(10):713-23.
Ideo H, Matsuzaka T, Nonaka T, Seko A, Yamashita K. Galectin-8-N-domain recognition mechanism for sialylated and sulfated glycans. J Biol Chem. 2011;286(13):11346- 55.
Johannes L. The Cellular and Chemical Biology of Endocytic Trafficking and Intracellular Delivery-The GL-Lect Hypothesis. Molecules. 2021;26(11).
Salomonsson E, Larumbe A, Tejler J, Tullberg E, Rydberg H, Sundin A, Khabut A, Frejd T, Lobsanov YD, Rini JM, Nilsson UJ, Leffler H. Monovalent interactions of galectin-1. Biochemistry. 2010;49(44):9518-32.
Lepur A, Salomonsson E, Nilsson UJ, Leffler H. Ligand induced galectin-3 protein self-association. J Biol Chem. 2012;287(26):21751-6.
Zhang Z, Miller MC, Xu X, Song C, Zhang F, Zheng Y, Zhou Y, Tai G, Mayo KH. NMR-based insight into galectin-3 binding to endothelial cell adhesion molecule CD146: Evidence for noncanonical interactions with the lectin's CRD beta-sandwich F-face. Glycobiology. 2019;29(8):608-18.
Mirgorodskaya E, Dransart E, Shafaq-Zadah M, Roderer D, Sihlbom C, Leffler H, Johannes L. Site-specific N-glycan profiles of alpha5 beta1 integrin from rat liver. Biol Cell. 2022;114(6):160-76.
Gouin SG, Garcia Fernandez JM, Vanquelef E, Dupradeau FY, Salomonsson E, Leffler H, Ortega-Munoz M, Nilsson UJ, Kovensky J. Multimeric lactoside "click clusters" as tools to investigate the effect of linker length in specific interactions with peanut lectin, galectin-1, and -3. Chembiochem. 2010;11(10):1430-42.
Stowell SR, Cho M, Feasley CL, Arthur CM, Song X, Colucci JK, Karmakar S, Mehta P, Dias-Baruffi M, McEver RP, Cummings RD. Ligand reduces galectin-1 sensitivity to oxidative inactivation by enhancing dimer formation. J Biol Chem. 2009;284(8):4989-99.
Hsu DK, Zuberi RI, Liu FT. Biochemical and biophysical characterization of human recombinant IgE-binding protein, an S-type animal lectin. J Biol Chem. 1992;267(20):14167-74.
Massa SM, Cooper DN, Leffler H, Barondes SH. L-29, an endogenous lectin, binds to glycoconjugate ligands with positive cooperativity. Biochemistry. 1993;32(1):260-7.
Sato S. Why does galectin-3 have a unique intrinsically disordered region? “Raison d’être” for the disordered structure and liquid–liquid phase separation –Part 2–. Glycoforum. 2023;26:A8: https://doi.org/10.32285/glycoforum.26A8J
Sato S. Why does galectin-3 have a unique intrinsically disordered region? “Raison d’être” for the disordered structure and liquid–liquid phase separation –Part 1–. Glycoforum. 2023;26:A1: https://doi.org/10.32285/glycoforum.26A1J
Leffler H. The life of a galectin: some questions about time, concentrations and affinities for its cellular and in vivo roles. Glycoforum. 2022;25:A12: https://doi.org/10.32285/glycoforum.25A12J
Carlsson S, Carlsson MC, Leffler H. Intracellular sorting of galectin-8 based on carbohydrate fine specificity. Glycobiology. 2007;17(9):906-12.
Lakshminarayan R, Wunder C, Becken U, Howes MT, Benzing C, Arumugam S, Sales S, Ariotti N, Chambon V, Lamaze C, Loew D, Shevchenko A, Gaus K, Parton RG, Johannes L. Galectin-3 drives glycosphingolipid-dependent biogenesis of clathrin- independent carriers. Nat Cell Biol. 2014;16(6):595-606.
Renard HF, Tyckaert F, Lo Giudice C, Hirsch T, Valades-Cruz CA, Lemaigre C, Shafaq-Zadah M, Wunder C, Wattiez R, Johannes L, van der Bruggen P, Alsteens D, Morsomme P. Endophilin-A3 and Galectin-8 control the clathrin-independent endocytosis of CD166. Nat Commun. 2020;11(1):1457.
Peterson K, Kumar R, Stenstrom O, Verma P, Verma PR, Hakansson M, Kahl- Knutsson B, Zetterberg F, Leffler H, Akke M, Logan DT, Nilsson UJ. Systematic Tuning of Fluoro-galectin-3 Interactions Provides Thiodigalactoside Derivatives with Single-Digit nM Affinity and High Selectivity. J Med Chem. 2018;61(3):1164-75.