Haik Mkhikian1, Michael Sy2, James W. Dennis3,4* and Michael Demetriou2*
1 Department of Pathology and Laboratory Medicine, University of California, Irvine, USA
2 Department of Neurology and Institute for Immunology, University of California, Irvine, CA 92697, USA.
3 Lunenfeld Tanenbaum Research Institute, Mount Sinai Hospital, 600 University Avenue R988, Toronto, Ontario, Canada M5G 1X5
4 Department of Molecular Genetics, & Department of Laboratory Medicine and Pathology, University of Toronto, Canada
* Corresponding authors:
James W. Dennis: DENNIS@lunenfeld.ca
Michael Demetriou: mdemetri@uci.edu
Haik Mkhikian
Dr. Haik Mkhikianは、2005年にカリフォルニア大学アーバイン校で哲学と生化学・分子生物学の学士号を取得した。さらに、メリーランド州ベセスダのNIHでUGSP奨学生として1年間過ごした後、カリフォルニア大学アーバイン校に戻ってMSTPのトレーニングを受けた。大学院では、Dr. Michael Demetriouの研究室で、T細胞におけるN型糖鎖分岐の制御と、多発性硬化症におけるその制御の異常について研究した。2016年にMD/PhDの研修を終えた後、カリフォルニア大学アーバイン校にリサーチトラックの臨床病理学レジデントとして残り、現在はリサーチフェローとして勤務している。
Michael Sy
Dr. Michael Syは、カリフォルニア大学アーバイン校神経学部門の助教であるとともに、バージニア州ロングビーチにある多発性硬化症地域センター(Multiple Sclerosis Regional Center)の共同ディレクターも勤めている。2002年にイェール大学を卒業した後、2011年にカリフォルニア大学アーバイン校のMedical Scientist Training ProgramでMDおよびPhDを取得した。さらに、UC Irvine Medical Centerにおいて、神経学のレジデント研修と神経免疫学のフェローシップ(2016年)を修了した。現在は、包括的かつホリスティックな患者ケアの提供に加えて、多発性硬化症のミエリン形成に関する基礎、橋渡し、および臨床研究を行っている。
James Dennis
Dr. James Dennisは、モントリオールのコンコルディア大学で数学の学士号を取得後、キングストンのクイーンズ大学で生化学の博士号を取得した(1978年)。ハイデルベルクのドイツがん研究センター(DKFZ)でポスドクとしてトレーニングを受けた後、Terry Foxフェローシップを獲得し、トロント大学のDr. Harry Schachterのもとで研究を行った(1980年)。クイーンズ大学の助教に就任した後(1982年)、ルーネンフェルド・タネンバウム研究所の創設メンバーとなり(1985年)、さらにトロント大学分子遺伝学部門の教授となった。カナダ王立協会のフェローであり、糖鎖生物学分野のCanada Research Chairに選出されている。20件の特許を保有しているほか、200以上の論文を発表し、被引用回数は20,000回に達している。また、多くの優秀な学生を指導していることも特筆に値する。
Michael Demetriou
Dr. Michael Demetriouは、トロント大学で修士号および博士号(1996年)を取得した(Dr. James Dennisの指導の下、分子遺伝学の博士号を取得)。トロント大学で神経科のレジデントを務めた後、ルーネンフェルド・タネンバウム研究所にいたDr. James Dennisのもとでポスドクとして糖鎖生物学のトレーニングを受け(2001年)、さらにカリフォルニア大学アーバイン校のDr. Stanley van den Noortのもとで神経免疫学の臨床フェローシップを修了した(2002年)。2001年には、カリフォルニア大学アーバイン校の教員となり、現在は神経学・微生物学・分子遺伝学分野の教授として、臨床・研究両面で活躍している。NIHから20年間継続して研究資金の提供を受けており、発表した44の論文は、5000回以上引用されている。また、NIHの2つの研究部会においてチャーターメンバーを務めたほか、これまでに9人の博士号取得者に加え、多数のポスドクおよび学部生を指導している。
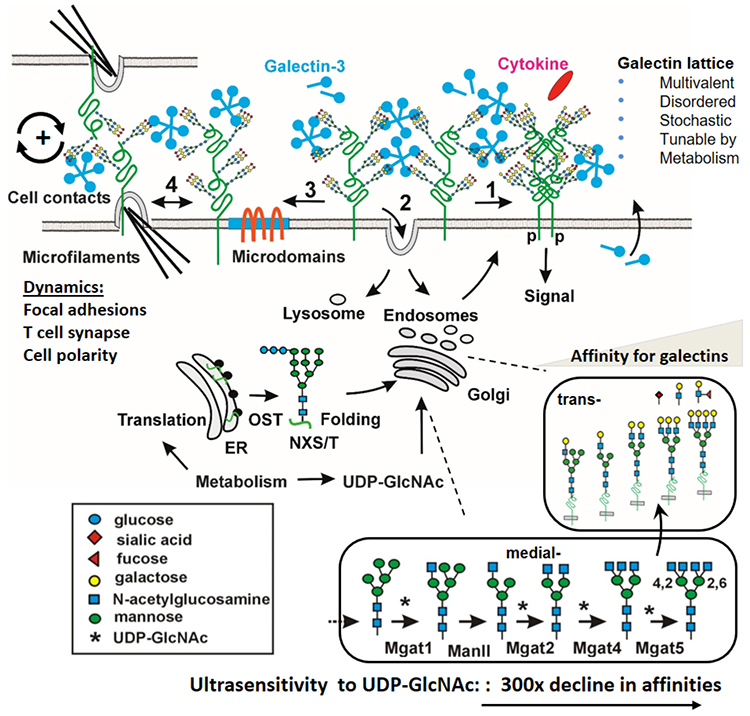
生細胞においては、ガレクチン格子は平面状の液-ドロップレット相であり、膜貫通型の糖タンパク質によって膜の近くに保持されている。多様な糖タンパク質から構成され、かつ柔軟な糖鎖構造によって、その配置は無定形状態である21,22(図 1)。Gal-3のCRDは、N-アセチルラクトサミン二糖単位(LacNAc、Gal β1-3/4GlcNAc)との結合に際し、配座エントロピーを増加するが、これは高親和性の受容体-リガンド間結合にはあまり見られないことである。より大きなCRDドメインのエントロピー移動が、結合したN型糖鎖におけるエントロピー低下を補うと考えられ、そのことが迅速なリガンドの結合・解離やリガンド交換に有利にはたらくのだろう23,24。Gal-3およびGal-9は糖タンパク質受容体の側方への拡散速度を低下させることが、FRAP(fluorescence recovery after photobleaching)法を用いた測定によって示されている25。例えば、Gal-9濃度やN型糖鎖の分岐、エンドサイトーシスの速度が変化した場合、初代肝細胞上にあるグルカゴン受容体のFRAP半減期も、エンドサイトーシスによる損失から受容体を保護するのと同じように変化した26。また、細胞表面におけるGal-3の多量体化がFRET(fluorescence resonance energy transfer)法によって測定されている他27、β1インテグリン受容体に結合したGal-3格子の動態が一粒子追跡法によって測定されている28。
このように、経口GlcNAc投与は、炎症性のT細胞およびB細胞応答を負に制御することと関連して、MSの病因および重症化を引き起こす4つの主要なメカニズム、すなわち、炎症性のT細胞応答、自然免疫における炎症性のB細胞活性、ミエリン修復の不全、および神経変性を抑制するユニークな手段といえる。このような多様な作用機序は、炎症反応は標的としているがミエリン修復や神経変性は標的としていない現在のMS治療薬にはないものであり、GlcNAcがMS患者の治療に有益であることを示唆している。この可能性を追求するため、Dr. Michael Demetriouが中心となり、MS患者に対するGlcNAc経口投与の非盲検臨床試験が、現在行われている。経口GlcNAcは忍容性が高いことが示されているほか、血清GlcNAcレベルを上昇させ、臨床的に有益な効果が認められた可能性がある。これらの知見を確認するために、追加の盲検試験が計画されている。
J.W.D.の研究は、カナダ・Canadian Institutes of Health Research(MOP-126183、MOP-136789、MOP-126029)の助成金およびCanada Research Chairs Programによって支援されている。M.D.の研究は、米国・National Institute of Allergy and Infectious Diseases(R01 AI108917)および米・National Center for Complementary and Integrative Health)(R01 AT007452)によって支援されている。
References
Ochieng, J., Platt, D., Tait, L., Hogan, V., Raz, T., Carmi, P., and Raz, A. (1993) Structure-function relationship of a recombinant human galactoside-binding protein. Biochemistry32, 4455-4460
Raz, A., Carmi, P., and Pazerini, G. (1988) Expression of two different endogenous galactoside-binding lectins sharing sequence homology. Cancer Res48, 645-649
Barondes, S. H., Cooper, D. N., Gitt, M. A., and Leffler, H. (1994) Galectins. Structure and function of a large family of animal lectins. J Biol Chem269, 20807-20810
Seetharaman, J., Kanigsberg, A., Slaaby, R., Leffler, H., Barondes, S. H., and Rini, J. M. (1998) X-ray crystal structure of the human galectin-3 carbohydrate recognition domain at 2.1-A resolution. J Biol Chem273, 13047-13052
Grigorian, A., and Demetriou, M. (2010) Manipulating cell surface glycoproteins by targeting N-glycan-galectin interactions. Methods Enzymol. 480, 245-266
Nabi, I. R., Shankar, J., and Dennis, J. W. (2015) The galectin lattice at a glance. J Cell Sci128, 2213-2219
Cooper, D. N. (2002) Galectinomics: finding themes in complexity. Biochim Biophys Acta1572, 209-231
Vasta, G. R. (2020) Galectins in Host-Pathogen Interactions: Structural, Functional and Evolutionary Aspects. Adv Exp Med Biol1204, 169-196
Thurston, T. L., Wandel, M. P., von Muhlinen, N., Foeglein, A., and Randow, F. (2012) Galectin 8 targets damaged vesicles for autophagy to defend cells against bacterial invasion. Nature482, 414-418
Kim, B. W., Hong, S. B., Kim, J. H., Kwon, D. H., and Song, H. K. (2013) Structural basis for recognition of autophagic receptor NDP52 by the sugar receptor galectin-8. Nat Commun4, 1613
Thurston, T. L., Boyle, K. B., Allen, M., Ravenhill, B. J., Karpiyevich, M., Bloor, S., Kaul, A., Noad, J., Foeglein, A., Matthews, S. A., Komander, D., Bycroft, M., and Randow, F. (2016) Recruitment of TBK1 to cytosol-invading Salmonella induces WIPI2-dependent antibacterial autophagy. EMBO J35, 1779-1792
Herrmann, J., Turck, C. W., Atchison, R. E., Huflejt, M. E., Poulter, L., Gitt, M. A., Burlingame, A. L., Barondes, S. H., and Leffler, H. (1993) Primary structure of the soluble lactose binding lectin L-29 from rat and dog and interaction of its non-collagenous proline-, glycine-, tyrosine-rich sequence with bacterial and tissue collagenase. J Biol Chem268, 26704-26711
Lin, Y. H., Qiu, D. C., Chang, W. H., Yeh, Y. Q., Jeng, U. S., Liu, F. T., and Huang, J. R. (2017) The intrinsically disordered N-terminal domain of galectin-3 dynamically mediates multisite self-association of the protein through fuzzy interactions. J Biol Chem292, 17845-17856
Ahmad, N., Gabius, H. J., Andre, S., Kaltner, H., Sabesan, S., Roy, R., Liu, B., Macaluso, F., and Brewer, C. F. (2003) Galectin-3 precipitates as a pentamer with synthetic multivalent carbohydrates and forms heterogeneous cross-linked complexes. J.Biol.Chem. 279, 10841-10847
Halimi, H., Rigato, A., Byrne, D., Ferracci, G., Sebban-Kreuzer, C., ElAntak, L., and Guerlesquin, F. (2014) Glycan dependence of Galectin-3 self-association properties. PLoS One9, e111836
Lepur, A., Salomonsson, E., Nilsson, U. J., and Leffler, H. (2012) Ligand induced galectin-3 protein self-association. J Biol Chem287, 21751-21756
Leppanen, A., Stowell, S., Blixt, O., and Cummings, R. D. (2005) Dimeric galectin-1 binds with high affinity to alpha2,3-sialylated and non-sialylated terminal N-acetyllactosamine units on surface-bound extended glycans. J Biol Chem280, 5549-5562
Dennis, J. W., and Brewer, C. F. (2013) Density-dependent lectin-glycan interactions as a paradigm for conditional regulation by posttranslational modifications. Mol Cell Proteomics12, 913-920
Demetriou, M., Granovsky, M., Quaggin, S., and Dennis, J. W. (2001) Negative regulation of T-cell activation and autoimmunity by Mgat5 N-glycosylation. Nature409, 733-739
Li, P., Banjade, S., Cheng, H. C., Kim, S., Chen, B., Guo, L., Llaguno, M., Hollingsworth, J. V., King, D. S., Banani, S. F., Russo, P. S., Jiang, Q. X., Nixon, B. T., and Rosen, M. K. (2012) Phase transitions in the assembly of multivalent signalling proteins. Nature483, 336-340
Dennis, J. W. (2015) Many Light Touches Convey the Message. Trends Biochem Sci40, 673-686
Guttman, M., Weinkam, P., Sali, A., and Lee, K. K. (2013) All-atom ensemble modeling to analyze small-angle x-ray scattering of glycosylated proteins. Structure21, 321-331
Diehl, C., Genheden, S., Modig, K., Ryde, U., and Akke, M. (2009) Conformational entropy changes upon lactose binding to the carbohydrate recognition domain of galectin-3. J Biomol NMR45, 157-169
Diehl, C., Engstrom, O., Delaine, T., Hakansson, M., Genheden, S., Modig, K., Leffler, H., Ryde, U., Nilsson, U. J., and Akke, M. (2010) Protein flexibility and conformational entropy in ligand design targeting the carbohydrate recognition domain of galectin-3. J Am Chem Soc132, 14577-14589
Lajoie, P., Partridge, E. A., Guay, G., Goetz, J. G., Pawling, J., Lagana, A., Joshi, B., Dennis, J. W., and Nabi, I. R. (2007) Plasma membrane domain organization regulates EGFR signaling in tumor cells. J Cell Biol179, 341-356
Johswich, A., Longuet, C., Pawling, J., Rahman, A. A., Ryczko, M., Drucker, D. J., and Dennis, J. W. (2014) N-Glycan Remodeling on Glucagon Receptor Is an Effector of Nutrient Sensing by the Hexosamine Biosynthesis Pathway. J Biol Chem289, 15927-15941
Nieminen, J., Kuno, A., Hirabayashi, J., and Sato, S. (2007) Visualization of galectin-3 oligomerization on the surface of neutrophils and endothelial cells using fluorescence resonance energy transfer. J Biol Chem282, 1374-1383
Yang, E. H., Rode, J., Howlader, M. A., Eckermann, M., Santos, J. T., Hernandez Armada, D., Zheng, R., Zou, C., and Cairo, C. W. (2017) Galectin-3 alters the lateral mobility and clustering of beta1-integrin receptors. PLoS One12, e0184378
Lagana, A., Goetz, J. G., Cheung, P., Raz, A., Dennis, J. W., and Nabi, I. R. (2006) Galectin binding to Mgat5-modified N-glycans regulates fibronectin matrix remodeling in tumor cells. Mol Cell Biol26, 3181-3193
Boscher, C., and Nabi, I. R. (2013) Galectin-3- and phospho-caveolin-1-dependent outside-in integrin signaling mediates the EGF motogenic response in mammary cancer cells. Mol Biol Cell24, 2134-2145
Snarr, B. D., St-Pierre, G., Ralph, B., Lehoux, M., Sato, Y., Rancourt, A., Takazono, T., Baistrocchi, S. R., Corsini, R., Cheng, M. P., Sugrue, M., Baden, L. R., Izumikawa, K., Mukae, H., Wingard, J. R., King, I. L., Divangahi, M., Satoh, M. S., Yipp, B. G., Sato, S., and Sheppard, D. C. (2020) Galectin-3 enhances neutrophil motility and extravasation into the airways during Aspergillus fumigatus infection. PLoS Pathog16, e1008741
Marhuenda, E., Fabre, C., Zhang, C., Martin-Fernandez, M., Iskratsch, T., Saleh, A., Bauchet, L., Cambedouzou, J., Hugnot, J. P., Duffau, H., Dennis, J. W., Cornu, D., and Bakalara, N. (2021) Glioma stem cells invasive phenotype at optimal stiffness is driven by MGAT5 dependent mechanosensing. J Exp Clin Cancer Res40, 139
Zick, Y., Eisenstein, M., Goren, R. A., Hadari, Y. R., Levy, Y., and Ronen, D. (2002) Role of galectin-8 as a modulator of cell adhesion and cell growth. Glycoconj J19, 517-526
Levy, Y., Arbel-Goren, R., Hadari, Y. R., Eshhar, S., Ronen, D., Elhanany, E., Geiger, B., and Zick, Y. (2001) Galectin-8 functions as a matricellular modulator of cell adhesion. J.Biol.Chem. 276, 31285-31295
Chen, I. J., Chen, H. L., and Demetriou, M. (2007) Lateral compartmentalization of T cell receptor versus CD45 by galectin-N-glycan binding and microfilaments coordinate basal and activation signaling. J Biol Chem282, 35361-35372
Demetriou, M., Granovsky, M., Quaggin, S., and Dennis, J. W. (2001) Negative regulation of T-cell activation and autoimmunity by Mgat5 N-glycosylation. Nature409, 733-739
Zhou, R. W., Mkhikian, H., Grigorian, A., Hong, A., Chen, D., Arakelyan, A., and Demetriou, M. (2014) N-glycosylation bidirectionally extends the boundaries of thymocyte positive selection by decoupling Lck from Ca(2)(+) signaling. Nat Immunol15, 1038-1045
Hirabayashi, J., Hashidate, T., Arata, Y., Nishi, N., Nakamura, T., Hirashima, M., Urashima, T., Oka, T., Futai, M., Muller, W. E., Yagi, F., and Kasai, K. (2002) Oligosaccharide specificity of galectins: a search by frontal affinity chromatography. Biochim.Biophys.Acta1572 232 -254
Lee, R. T., and Lee, Y. C. (2000) Affinity enhancement by multivalent lectin-carbohydrate interaction. Glycoconj J17, 543-551
Dam, T. K., and Brewer, C. F. (2008) Effects of clustered epitopes in multivalent ligand-receptor interactions. Biochemistry47, 8470-8476
Patnaik, S. K., Potvin, B., Carlsson, S., Sturm, D., Leffler, H., and Stanley, P. (2006) Complex N-glycans are the major ligands for galectin-1, -3, and -8 on Chinese hamster ovary cells. Glycobiology16, 305-317
Do, K.-Y., Fregien, N., Pierce, M., and Cummings, R. D. (1994) Modification of glycoproteins by N-acetylglucosaminyltransferase V is greatly influenced by accessibility of the enzyme to oligosacharide acceptors. J.Biol.Chem. 269, 23456-23464
Losfeld, M. E., Scibona, E., Lin, C. W., Villiger, T. K., Gauss, R., Morbidelli, M., and Aebi, M. (2017) Influence of protein/glycan interaction on site-specific glycan heterogeneity. FASEB J31, 4623-4635
Yu, R., Longo, J., van Leeuwen, J. E., Zhang, C., Branchard, E., Elbaz, M., Cescon, D. W., Drake, R. R., Dennis, J. W., and Penn, L. Z. (2021) Mevalonate pathway inhibition slows breast cancer metastasis via reduced N-glycosylation abundance and branching. Cancer Res
Lau, K. S., Partridge, E. A., Grigorian, A., Silvescu, C. I., Reinhold, V. N., Demetriou, M., and Dennis, J. W. (2007) Complex N-glycan number and degree of branching cooperate to regulate cell proliferation and differentiation. Cell129, 123-134
Varki, A. (2011) Evolutionary forces shaping the Golgi glycosylation machinery: why cell surface glycans are universal to living cells. Cold Spring Harb Perspect Biol3
Schachter, H. (1986) Biosynthetic controls that determine the branching and microheterogeneity of protein-bound oligosaccharides. Biochem.Cell Biol. 64, 163-181
Sasai, K., Ikeda, Y., Fujii, T., Tsuda, T., and Taniguchi, N. (2002) UDP-GlcNAc concentration is an important factor in the biosynthesis of beta1,6-branched oligosaccharides: regulation based on the kinetic properties of N-acetylglucosaminyltransferase V. Glycobiology12, 119 -127
Hassinen, A., and Kellokumpu, S. (2014) Organizational interplay of Golgi N-glycosyltransferases involves organelle microenvironment-dependent transitions between enzyme homo- and heteromers. J Biol Chem289, 26937-26948
Climer, L. K., Dobretsov, M., and Lupashin, V. (2015) Defects in the COG complex and COG-related trafficking regulators affect neuronal Golgi function. Front Neurosci9, 405
Zhen, Y., Caprioli, R. M., and Staros, J. V. (2003) Characterization of glycosylation sites of the epidermal growth factor receptor. Biochemistry42, 5478-5492
Valiant, L. (2013) Probably Approximately Correct: Nature's Algorithms for Learning and Prospering in a Complex World, Basic Books, Inc. , New York, NY
Zhang, F., Wang, S., Yin, L., Yang, Y., Guan, Y., Wang, W., Xu, H., and Tao, N. (2015) Quantification of epidermal growth factor receptor expression level and binding kinetics on cell surfaces by surface plasmon resonance imaging. Anal Chem87, 9960-9965
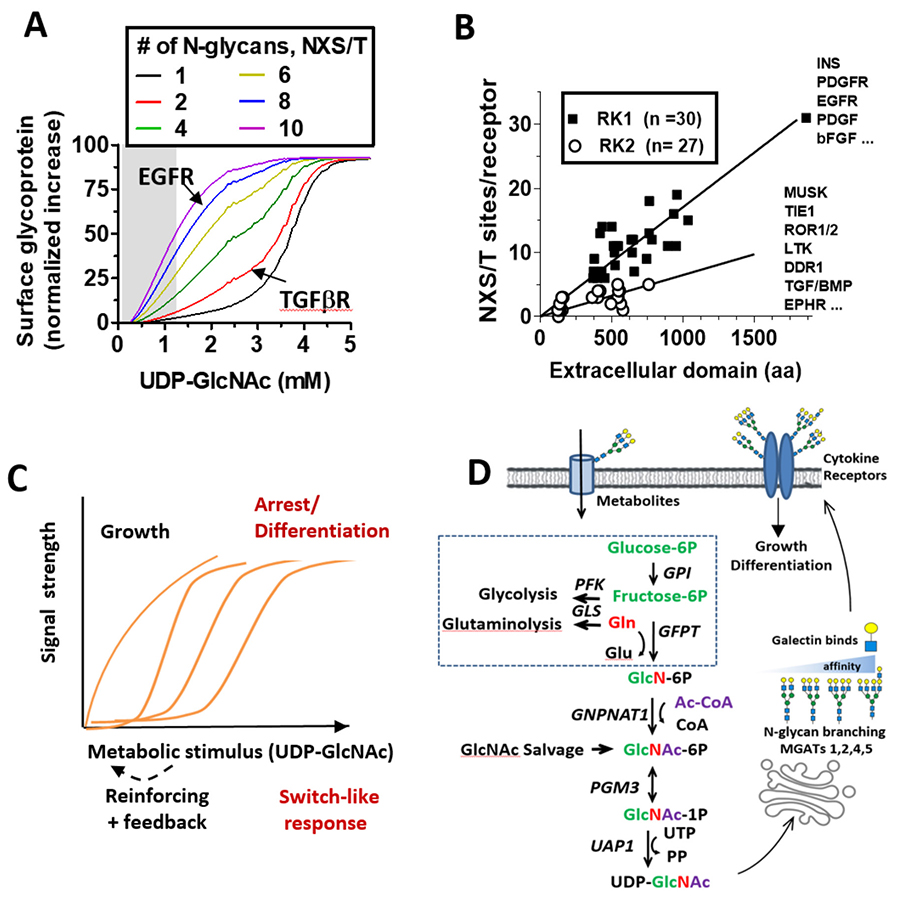
Araujo, L., Khim, P., Mkhikian, H., Mortales, C. L., and Demetriou, M. (2017) Glycolysis and glutaminolysis cooperatively control T cell function by limiting metabolite supply to N-glycosylation. Elife6
Grigorian, A., Lee, S. U., Tian, W., Chen, I. J., Gao, G., Mendelsohn, R., Dennis, J. W., and Demetriou, M. (2007) Control of T Cell-mediated autoimmunity by metabolite flux to N-glycan biosynthesis. J Biol Chem282, 20027-20035
Mkhikian, H., Grigorian, A., Li, C. F., Chen, H. L., Newton, B., Zhou, R. W., Beeton, C., Torossian, S., Tatarian, G. G., Lee, S. U., Lau, K., Walker, E., Siminovitch, K. A., Chandy, K. G., Yu, Z., Dennis, J. W., and Demetriou, M. (2011) Genetics and the environment converge to dysregulate N-glycosylation in multiple sclerosis. Nat Commun2, 334
Ohtsubo, K., Takamatsu, S., Gao, C., Korekane, H., Kurosawa, T. M., and Taniguchi, N. (2013) N-Glycosylation modulates the membrane sub-domain distribution and activity of glucose transporter 2 in pancreatic beta cells. Biochem Biophys Res Commun434, 346-351
Ohtsubo, K., Takamatsu, S., Minowa, M. T., Yoshida, A., Takeuchi, M., and Marth, J. D. (2005) Dietary and genetic control of glucose transporter 2 glycosylation promotes insulin secretion in suppressing diabetes. Cell123, 1307-1321
Takamatsu, S., Oguri, S., Minowa, M. T., Yoshida, A., Nakamura, K., Takeuchi, M., and Kobata, A. (1999) Unusually high expression of N-acetylglucosaminyltransferase-IV a in human choriocarcinoma cell lines: a possible enzymatic basis of the formaiton of abnormal biantennary sugar chain. Cancer Research59, 3949-3953
Ohno, M., Nishikawa, A., Koketsu, M., Taga, H., Endo, Y., Hada, T., Higashino, K., and Taniguchi, N. (1992) Enzymatic basis of sugar structures of alpha-fetoprotein in hepatoma and hepatoblastoma cell lines: correlation with activities of alpha 1-6 fucosyltransferase and N-acetylglucosaminyltransferases III and V. Int.J.Cancer51, 315-317
Lau, K. S., and Dennis, J. W. (2008) N-Glycans in cancer progression. Glycobiology18, 750-760
Ryczko, M. C., Pawling, J., Chen, R., Abdel Rahman, A. M., Yau, K., Copeland, J. K., Zhang, C., Surendra, A., Guttman, D. S., Figeys, D., and Dennis, J. W. (2016) Metabolic Reprogramming by Hexosamine Biosynthetic and Golgi N-Glycan Branching Pathways. Sci Rep6, 23043
Dennis, J. W. (2017) Genetic code asymmetry supports diversity through experimentation with posttranslational modifications. Curr Opin Chem Biol41, 1-11
Lee, S. U., Grigorian, A., Pawling, J., Chen, I. J., Gao, G., Mozaffar, T., McKerlie, C., and Demetriou, M. (2007) N-glycan processing deficiency promotes spontaneous inflammatory demyelination and neurodegeneration. J Biol Chem282, 33725-33734
Grigorian, A., Araujo, L., Naidu, N. N., Place, D., Choudhury, B., and Demetriou, M. (2011) N-acetylglucosamine inhibits T-helper 1 (Th1) / T-helper 17 (Th17) responses and treats experimental autoimmune encephalomyelitis. J. Biol. Chem. 286, 40133-40141
Grigorian, A., Lee, S. U., Tian, W., Chen, I. J., Gao, G., Mendelsohn, R., Dennis, J. W., and Demetriou, M. (2007) Control of T Cell-mediated autoimmunity by metabolite flux to N-glycan biosynthesis. J Biol Chem282, 20027-20035
Morgan, R., Gao, G., Pawling, J., Dennis, J. W., Demetriou, M., and Li, B. (2004) N-acetylglucosaminyltransferase V (Mgat5)-mediated N-glycosylation negatively regulates Th1 cytokine production by T cells. J Immunol173, 7200-7208
Mortales, C. L., Lee, S. U., and Demetriou, M. (2020) N-Glycan Branching Is Required for Development of Mature B Cells. J Immunol205, 630-636
Mortales, C. L., Lee, S. U., Manousadjian, A., Hayama, K. L., and Demetriou, M. (2020) N-Glycan Branching Decouples B Cell Innate and Adaptive Immunity to Control Inflammatory Demyelination. iScience23, 101380
Li, C. F., Zhou, R. W., Mkhikian, H., Newton, B. L., Yu, Z., and Demetriou, M. (2013) Hypomorphic MGAT5 polymorphisms promote multiple sclerosis cooperatively with MGAT1 and interleukin-2 and 7 receptor variants. J Neuroimmunol256, 71-76
Backer-Koduah, P., Infante-Duarte, C., Ivaldi, F., Uccelli, A., Bellmann-Strobl, J., Wernecke, K. D., Sy, M., Demetriou, M., Dorr, J., Paul, F., and Ulrich Brandt, A. (2020) Effect of vitamin D supplementation on N-glycan branching and cellular immunophenotypes in MS. Ann Clin Transl Neurol7, 1628-1641
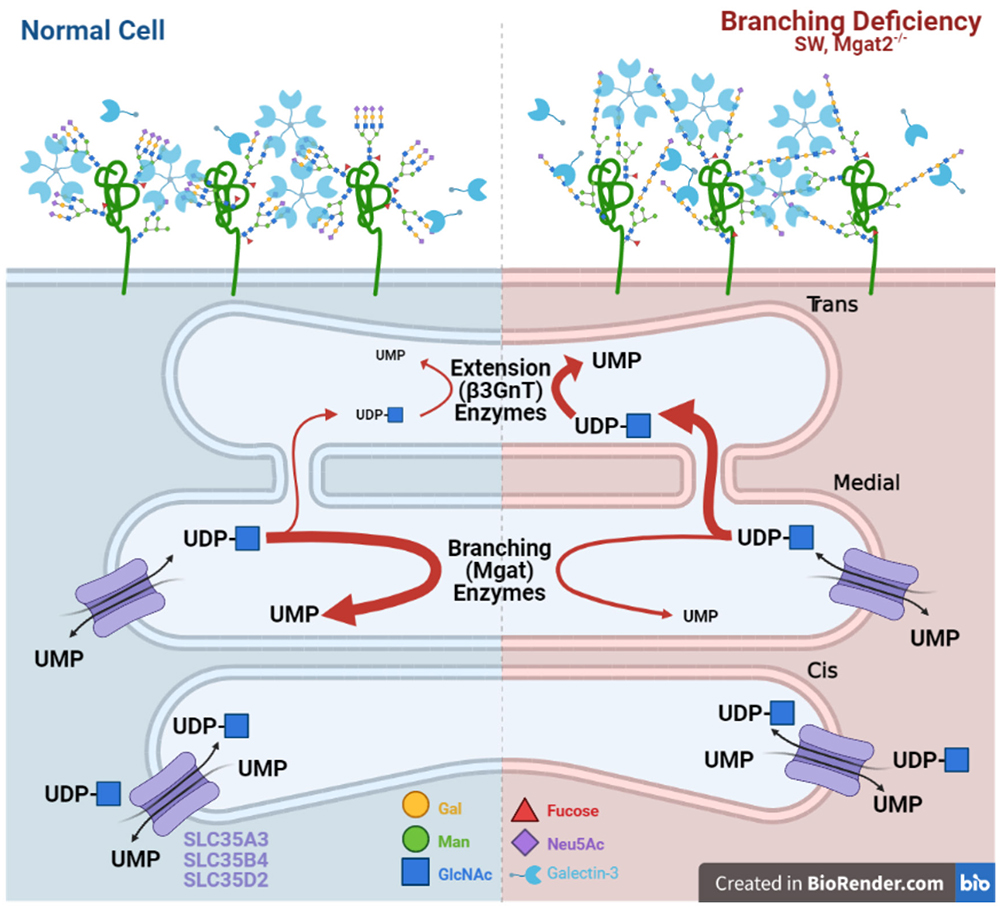
Mkhikian, H., Mortales, C. L., Zhou, R. W., Khachikyan, K., Wu, G., Haslam, S. M., Kavarian, P., Dell, A., and Demetriou, M. (2016) Golgi self-correction generates bioequivalent glycans to preserve cellular homeostasis. Elife5
Chui, D., Oh-Eda, M., Liao, Y.-F., Panneerselvam, K., Lal, A., Marek, K. W., Freeze, H. H., Moremen, K. W., Fukuda, M. N., and Marth, J. D. (1997) Alpha-mannosidase-II deficiency results in dyserythropoiesis and unveils an alternate pathway in oligosaccharide biosynthesis. Cell90, 157-167
Granovsky, M., Fata, J., Pawling, J., Muller, W. J., Khokha, R., and Dennis, J. W. (2000) Suppression of tumor growth and metastasis in Mgat5-deficient mice. Nat Med6, 306-312
Bell, T. J., Brand, O. J., Morgan, D. J., Salek-Ardakani, S., Jagger, C., Fujimori, T., Cholewa, L., Tilakaratna, V., Ostling, J., Thomas, M., Day, A. J., Snelgrove, R. J., and Hussell, T. (2019) Defective lung function following influenza virus is due to prolonged, reversible hyaluronan synthesis. Matrix Biol80, 14-28
Sy, M., Brandt, A. U., Lee, S. U., Newton, B. L., Pawling, J., Golzar, A., Rahman, A. M. A., Yu, Z., Cooper, G., Scheel, M., Paul, F., Dennis, J. W., and Demetriou, M. (2020) N-acetylglucosamine drives myelination by triggering oligodendrocyte precursor cell differentiation. J Biol Chem295, 17413-17424
Pasquini, L. A., Millet, V., Hoyos, H. C., Giannoni, J. P., Croci, D. O., Marder, M., Liu, F. T., Rabinovich, G. A., and Pasquini, J. M. (2011) Galectin-3 drives oligodendrocyte differentiation to control myelin integrity and function. Cell Death Differ18, 1746-1756
Zhang, Y., Yu, X., Ichikawa, M., Lyons, J. J., Datta, S., Lamborn, I. T., Jing, H., Kim, E. S., Biancalana, M., Wolfe, L. A., DiMaggio, T., Matthews, H. F., Kranick, S. M., Stone, K. D., Holland, S. M., Reich, D. S., Hughes, J. D., Mehmet, H., McElwee, J., Freeman, A. F., Freeze, H. H., Su, H. C., and Milner, J. D. (2014) Autosomal recessive phosphoglucomutase 3 (PGM3) mutations link glycosylation defects to atopy, immune deficiency, autoimmunity, and neurocognitive impairment. J Allergy Clin Immunol133, 1400-1409, 1409 e1401-1405
Miller, J. B., Bull, S., Miller, J., and McVeagh, P. (1994) The oligosaccharide composition of human milk: temporal and individual variations in monosaccharide components. J Pediatr Gastroenterol Nutr19, 371-376
Lawson, M. A. E., O'Neill, I. J., Kujawska, M., Gowrinadh Javvadi, S., Wijeyesekera, A., Flegg, Z., Chalklen, L., and Hall, L. J. (2020) Breast milk-derived human milk oligosaccharides promote Bifidobacterium interactions within a single ecosystem. ISME J14, 635-648
Brandt, A. U., Sy, M., Bellmann-Strobl, J., Newton, B. L., Pawling, J., Zimmermann, H. G., Yu, Z., Chien, C., Dorr, J., Wuerfel, J. T., Dennis, J. W., Paul, F., and Demetriou, M. (2021) Association of a Marker of N-Acetylglucosamine With Progressive Multiple Sclerosis and Neurodegeneration. JAMA Neurol