Barondes SH, Castronovo V, Cooper DN, Cummings RD, Drickamer K, Feizi T, Gitt MA, Hirabayashi J, Hughes C, Kasai K, Leffler H, Liu F-T, Lotan R, Mercurio AM, Monsigny M, Pillai S, Poirer F, Raz A, Rigby PWJ, Wang JL (1994) Galectins: a family of animal beta-galactoside-binding lectins. Cell. 76, 597–598.
Hirabayashi J, Hashidate T, Arata Y, Nishi N, Nakamura T, Hirashima M, Urashima T, Oka T, Futai M, Muller WEG, Yagi F, Kasai K (2002) Oligosaccharide specificity of galectins: a search by frontal affinity chromatography. Biochim Biophys Acta. 1572 (2–3), 232–254.
Iwaki J and Hirabayashi J (2018) Carbohydrate-binding specificity of human galectins: an overview by frontal affinity chromatography. Trends Glycosci Glycotechnol. 30(172), SE137-SE153.
Kasai K (1997) Galectin: intelligent glue, non-bureaucratic bureaucrat or almighty supporting actor. Trends Glycosci Glycotechnol. 9(45), 167–170.
Kobayashi Y, Tateno H, Ogawa H, Yamamoto K, Hirabayashi J (2014) Comprehensive list of lectins: origins, natures, and carbohydrate specificities. Methods Mol Biol. 1200, 555–577.
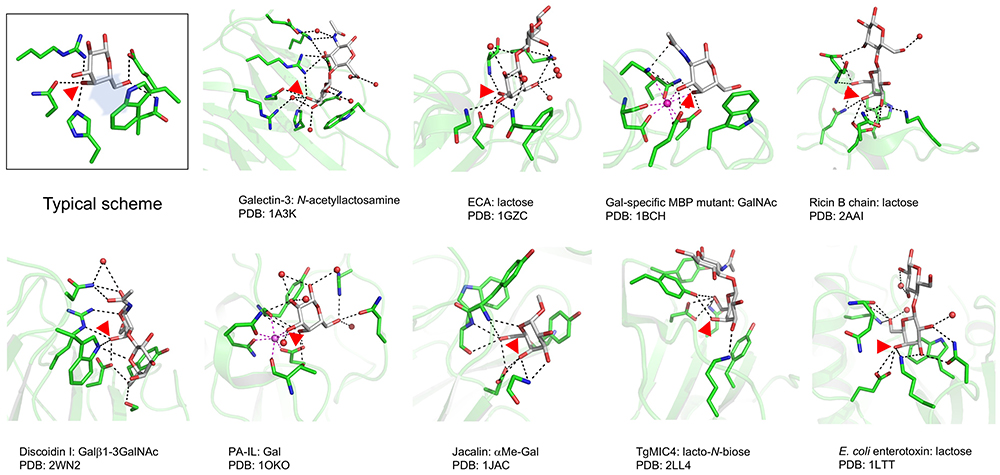
Iwaki J, Tateno H, Nishi N, Minamisawa T, Nakamura-Tsuruta S, Itakura Y, Kominami J, Urashima T, Nakamura T, Hirabayashi J (2011) The Galβ-(syn)-gauche configuration is required for galectin-recognition disaccharides. Biochim Biophys Acta. 1810(7), 643–651.
Rini JM (1995) Lectin structure. Annu Rev Biophys Biomol Struct. 24, 551–577.
Ozeki Y, Matsui T, Nitta K, Kawauchi H, Takayanagi Y, Titani K (1991) Purification and characterization of beta-galactoside binding lectin from frog (Rana catesbeiana) eggs. Biochem Biophys Res Commun. 178, 407–413.
Spurlino JC, Lu GY, Quiocho FA (1991) The 2.3-A resolution structure of the maltose- or maltodextrin-binding protein, a primary receptor of bacterial active transport and chemotaxis. J Biol Chem. 266(8), 5202–5219.
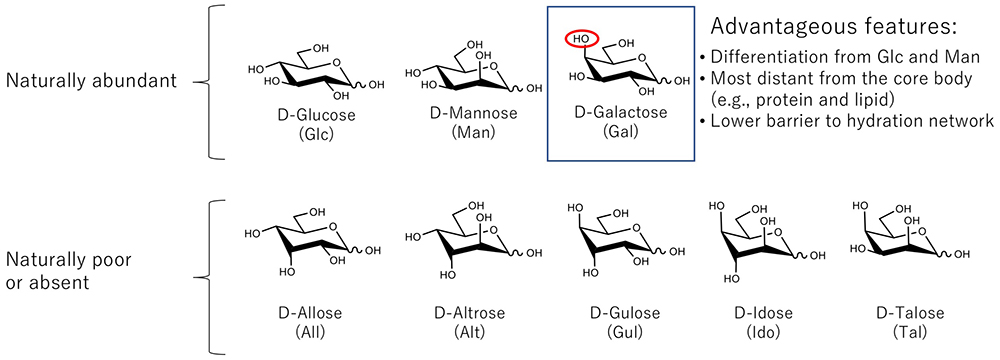
Hirabayashi J (1996) On the origin of elementary hexoses. Q Rev Biol. 71(3), 365–380.
Hirabayashi J (2001) Glycan structures, clues to the origin of saccharides. Viva origino. 29, 119–133.
Uedaira H, Okouchi S, Tsuda S, Uedaira H (2001) Hydration of glucose and galactose derivatives: hydration of sugar derivatives. Bull Chem Soc Jpn. 74, 1857–1861.
Hirabayashi J, Kasai K (1993) The family of metazoan metal-independent beta-galactoside-binding lectins: structure, function and molecular evolution. Glycobiology. 3(4), 297–304.
Cooper DN (2002) Galectinomics: finding themes in complexity. Biochim Biophys Acta. 1572(2–3), 209–231.
Gabius H-J (2021) Galectins: (much) more than ga(lactose-binding)lectins. Glycoforum. 24(1), A1.
Drickamer K (1988) Two distinct classes of carbohydrate-recognition domains in animal lectins. J Biol Chem. 263(20), 9557–9560.
Drickamer K, Dodd RB (1999) C-Type lectin-like domains in Caenorhabditis elegans: predictions from the complete genome sequence. Glycobiology. 9(12), 1357–1369.
Takeuchi T, Sennari R, Sugiura K, Tateno H, Hirabayashi J, Kasai K (2008) A C-type lectin of Caenorhabditis elegans: its sugar-binding property revealed by glycoconjugate microarray analysis. Biochem Biophys Res Commun. 377(1), 303–306.
Nemoto-Sasaki Y, Hayama K, Ohya H, Arata Y, Kaneko MK, Saitou N, Hirabayashi J, Kasai K (2008) Caenorhabditis elegans galectins LEC-1-LEC-11: structural features and sugar-binding properties. Biochim Biophys Acta. 1780(10), 1131–1142.
Pfeifer K, Haasemann M, Gamulin V, Bretting H, Fahrenholz F, Müller WE (1993) S-type lectins occur also in invertebrates: high conservation of the carbohydrate recognition domain in the lectin genes from the marine sponge Geodia cydonium. Glycobiology. 3(2), 179–184.
Vasta GR (2012) Galectins as pattern recognition receptors: structure, function, and evolution. Adv Exp Med Biol. 946, 21–36.
Vasta GR (2020) Galectins in host-pathogen interactions: structural, functional and evolutionary aspects. Adv Exp Med Biol. 1204,169–196.
Verkerke H, Dias-Baruffi M, Cummings RD, Arthur CM, Stowell SR (2022) Galectins: An ancient family of carbohydrate binding proteins with modern functions. Methods Mol Biol. 2442, 1–40.
Hirabayashi J, Kawasaki H, Suzuki K, Kasai K (1987) Complete amino acid sequence of 14 kDa beta-galactoside-binding lectin of chick embryo. J Biochem. 101(3), 775–787.
Sakakura Y, Hirabayashi J, Oda Y, Ohyama Y, Kasai K (1990) Structure of chicken 16-kDa beta-galactoside-binding lectin. Complete amino acid sequence, cloning of cDNA, and production of recombinant lectin. J Biol Chem. 265(35), 21573–21579.
Tsunasawa S, Stewart JW, Sherman F (1985) Amino-terminal processing of mutant forms of yeast iso-1-cytochrome c. The specificities of methionine aminopeptidase and acetyltransferase. J Biol Chem. 260(9), 5382–5391.
Ohyama Y, Hirabayashi J, Oda Y, Ohno S, Kawasaki H, Suzuki K, Kasai K (1986) Nucleotide sequence of chick 14K beta-galactoside-binding lectin mRNA. Biochem Biophys Res Commun. 134(1), 51–56.
Cooper DN, Barondes SH (1990) Evidence for export of a muscle lectin from cytosol to extracellular matrix and for a novel secretory mechanism. J Cell Biol. 110(5), 1681–1691.
Bänfer S, Jacob R. (2020) Galectins in intra- and extracellular vesicles. Biomolecules. 10(9), 1232.
Hong MH, Weng IC, Li FY, Lin WH, Liu FT (2021) Intracellular galectins sense cytosolically exposed glycans as danger and mediate cellular responses. J Biomed Sci. 28(1), 16.
La Venuta G, Zeitler M, Steringer JP, Müller HM, Nickel W (2015) The startling properties of fibroblast growth factor 2: how to exit mammalian cells without a signal peptide at hand. J Biol Chem. 290(45), 27015–27020.
Steringer JP, Nickel W (2018) A direct gateway into the extracellular space: Unconventional secretion of FGF2 through self-sustained plasma membrane pores. Semin Cell Dev Biol. 83, 3–7.
Rubartelli A, Cozzolino F, Talio M, Sitia R (1990) A novel secretory pathway for interleukin-1 beta, a protein lacking a signal sequence. EMBO J. 9(5), 1503–1510.
Popa SJ, Stewart SE, Moreau K (2018) Unconventional secretion of annexins and galectins. Semin Cell Dev Biol. 83, 42–50.
Dennis JW (2015) Many light touches convey the message. Trends Biochem Sci. 40(11), 673–686.
Dennis JW (2017) Genetic code asymmetry supports diversity through experimentation with posttranslational modifications. Curr Opin Chem Biol. 41, 1–11.
Demetriou M, Nabi IR, Dennis JW (2018) Galectins as adaptors: linking glycosylation and metabolism with extracellular cues. Trends Glycosci Glycotechnol. 30(172), SE167–SE177.