氏名:佐藤 祥子
ラヴァル大学医学部・医学部付属研究所、薬学博士
千葉大学薬学部卒業。1987年より東京大学医科学研究所大学院で木幡教授の指導を受ける。その間ロンドンの国立医学研究所にてR. Colin Hughes博士の下でガレクチン-3研究行う。1994年東京大学博士号取得。ついでスタンフォード大学のDr. Ron Kopito研究室でポスドク研究員として、遺伝性嚢胞性線維症への化学療法に関する研究を行った。1999年よりカナダ・ラヴァル大学医学部感染症研究センター糖質生物学研究室主任研究員、ラヴァル大学医学部准教授。2010年同大学同部教授。2003年以降同部門Bioimaging platformの長兼任。
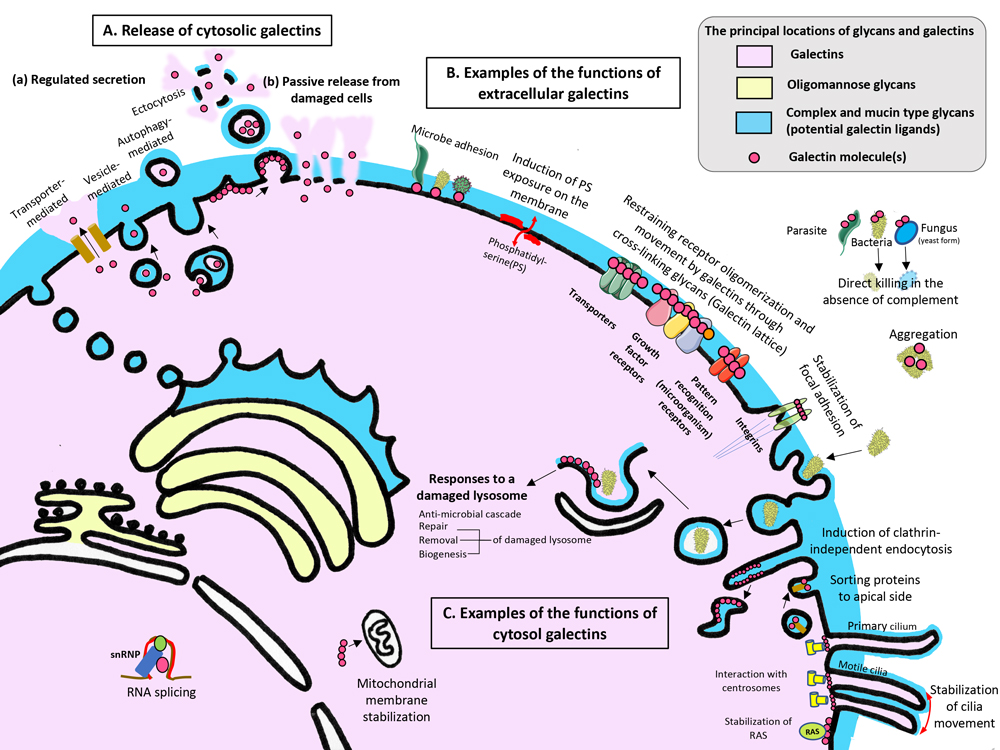
A. 細胞質に存在するガレクチンは、制御分泌経路(a)及び損傷した細胞からの受動的な放出(b)により細胞外環境に放出される。制御分泌経路の場合は、ガレクチンは4つの経路、すなわちトランスポーター、ベシクル、およびオートファジーを介した経路、またはエクトサイトーシスを介した経路により分泌されることが知られている。
B. 細胞外環境に放出されたガレクチンは、時空間的かつ状況依存的に多くの機能を発揮する。それらの機能のほとんどは、ガレクチンの糖鎖結合活性とその結合多価性(multivalency)に依存している。この図にはこれらの機能の一部を記した。
C. 細胞質内においては、ガレクチンは糖鎖−ガレクチンとタンパク質−ガレクチン相互作用を介してさまざまな応答を仲介する。 これらの機能の一部を記した。
詳細な説明については、佐藤によるグライコフォーラムの記事または論文を参照(S. Sato: Mammalian cytosolic galectins act as damage-associated molecular patterns, resolutions-associated molecular patterns, and pattern-recognition receptors in innate immunity, Comprehensive Glycoscience 2nd Edition, Elsevier, in press)。
■ Kenichi Kasai (Teikyo University, Japan)
■ Hans-Joachim Gabius (Ludwig-Maximilians-University Munich, Germany)
■ Hakon Leffler (Lund University, Sweden)
■ James W. Dennis (Mount Sinai Hospital, Toronto, Canada)
■ Tadasu Urashima (Obihiro University of Agriculture and Veterinary Medicine, Obihiro, Japan)
■ Sachiko Sato (Laval University, Quebec City, Canada)
■ Jun Hirabayashi (AIST, Tsukuba, Japan)
References
Barondes SH, Castronovo V, Cooper DN, Cummings RD, Drickamer K, Feizi T, Gitt MA, Hirabayashi J, Hughes C, Kasai K, Leffler H, Liu F-T, Lotan R, Mercurio AM, Monsigny M, Pillai S, Poirer F, Raz A, Rigby PWJ, Wang JL (1994) Galectins: a family of animal beta-galactoside-binding lectins. Cell. 76, 597–8.
Sato S (2020) Galectins and innate immunity, Comprehensive Glycoscience 2nd edition Vol 5 Chapter 147. In press
Kasai K (1997) Galectin: intelligent glue, non-bureaucratic bureaucrat or almighty supporting actor. Trends Glycosic Glycotechnol. 9(45), 167-70.
Nishi N, Abe A, Iwaki J, Yoshida H, Itoh A, Shoji H, Kamitori S, Hirabayashi J, Nakamura T (2008) Functional and structural bases of a cysteine-less mutant as a long-lasting substitute for galectin-1. Glycobiology. 18(12), 1065-73.
Nishi N, Itoh A, Fujiyama A, Yoshida N, Araya S, Hirashima M, Shoji H, Nakamura T (2005) Development of highly stable galectins: truncation of the linker peptide confers protease-resistance on tandem-repeat type galectins. FEBS Lett. 579(10), 2058-64.
Kilpatrick DC (2002) Animal lectins: a historical introduction and overview. Biochim Biophys Acta. 1572, 187–97.
Barondes SH (1997) Galectins: A personal overview. Trends Glycosci Glycotechnol. 9, 1–7.
Reitherman R W, Rosen SD, Frasier WA, Barondes SH (1975) Cell surface species-specific high affinity receptors for discoidin: developmental regulation in Dictyostelium discoideum. Proc Natl Acad Sci U S A. 72, 3541–5.
Teichberg VI, Silman I, Beitsch DD, Resheff G (1975) A beta-D-galactoside binding protein from electric organ tissue of Electrophorus electricus. Proc Natl Acad Sci U S A. 72, 1383–7.
Leffler H, Barondes SH (1986) Specificity of binding of three soluble rat lung lectins to substituted and unsubstituted mammalian b-galactosides. J Biol Chem. 261, 10119–26.
Ohyama Y, Hirabayashi J, Oda Y, Ohno S, Kawasaki H, Suzuki K, Kasai K (1986) Nucleotide sequence of chick 14K beta-galactoside-binding lectin mRNA. Biochem Biophys Res Commun. 134, 51–6.
Drickamer K (1988) Two distinct classes of carbohydrate-recognition domains in animal lectins. J Biol Chem. 263, 9557–60.
Cooper DN, Barondes SH (1990) Evidence for export of a muscle lectin from cytosol to extracellular matrix and for a novel secretory mechanism. J Cell Biol. 110, 1681–91.
Hirabayashi J, Satoh M, Kasai K (1992) Evidence that Caenorhabditis elegans 32-kDa beta-galactoside-binding protein is homologous to vertebrate beta-galactoside-binding lectins. cDNA cloning and deduced amino acid sequence. J Biol Chem. 267, 15485–90.
Hirabayashi J, Kasai K (1993) The family of metazoan metal-independent beta-galactoside-binding lectins: structure, function and molecular evolution. Glycobiology. 3, 297–304.
Lobsanov YD, Gitt MA, Leffler H, Barondes SH, Rini JM (1993) X-ray crystal structure of the human dimeric S-Lac lectin, L-14-II, in complex with lactose at 2.9-A resolution. J Biol Chem. 268, 27034–8.
Poirier F, Robertson EJ (1993) Normal development of mice carrying a null mutation in the gene encoding the L14 S-type lectin. Development. 119, 1229–36.
Barondes SH, Castronovo V, Cooper DN, Cummings RD, Drickamer K, Feizi T, Gitt MA, Hirabayashi J, Hughes C, Kasai K et al (1994) Galectins: a family of animal beta-galactoside-binding lectins. Cell. 76, 597–8.
Dagher SF, Wang JL, Patterson RJ (1995) Identification of galectin-3 as a factor in pre-mRAN splicing. Proc Natl Acad Sci U S A. 92, 1213–1217
Cooper DN, Boulianne RP, Charlton S, Farrell EM, Sucher A, Lu BC (1997) Fungal galectins, sequence and specificity of two isolectins from Coprinus cinereus. J Biol Chem. 272, 1514–21.
Matsumoto R, Matsumoto H, Seki M, Hata M, Asano Y, Kanegasaki S, Stevens RL, Hirashima M (1998) Human ecalectin, a variant of human galectin-9, is a novel eosinophil chemoattractant produced by T lymphocytes. J Biol Chem. 273, 16976–84.
Cooper DN, Barondes SH (1999) God must love galectins; he made so many of them. Glycobiology. 9, 979–84.
Hughes RC (1999) Secretion of the galectin family of mammalian carbohydrate-binding proteins. Biochim Biophys Acta. 1473, 172–85.
Demetriou M, Granovsky M, Quaggin S, Dennis JW (2001) Negative regulation of T-cell activation and autoimmunity by Mgat5 N-glycosylation. Nature. 409, 733–9.
Hirabayashi J, Hashidate, T, Arata Y, Nishi N, Nakamura T, Hirashima M, Urashima T, Oka T, Futai M, Muller WE, YagiF, Kasai K. (2002) Oligosaccharide specificity of galectins: a search by frontal affinity chromatography. Biochim Biophys Acta. 1572, 232–54.
Sasai K, Ikeda Y, Fujii T, Tsuda T, Taniguchi N (2002) UDP-GlcNAc concentration is an important factor in the biosynthesis of beta1,6-branched oligosaccharides: regulation based on the kinetic properties of N-acetylglucosaminyltransferase V. Glycobiology. 12, 119–27.
Sato S (2002) Galectin as a molecule of danger signal, which could evoke immune response to infection. Trends Glycosci Glycotechnol. 14, 285–301
Sato S, Nieminen J (2004) Seeing strangers or announcing “danger”: Galectin-3 in two models of innate immunity. Glycoconj J. 19, 583–91.
Delacour D, Gouyer V, Zanetta JP, Drobecq H, Leteurtre E, Grard G, Moreau-Hannedouche O, Maes E, Pons A, Andre S, Le Bivic A, Gabius HJ, Manninen A, Simons K, Huet G (2005) Galectin-4 and sulfatides in apical membrane trafficking in enterocyte-like cells. J Cell Biol. 169, 491–501
Kohatsu L, Hsu DK, Jegalian AG, Liu FT, Baum LG (2006) Galectin-3 induces death of Candida species expressing specific beta-1,2-linked mannans. J Immunol. 177, 4718–26.
Stowell SR, Karmakar S, Stowell CJ, Dias-Baruffi M, McEver RP, Cummings RD (2007) Human galectin-1, -2, and -4 induce surface exposure of phosphatidylserine in activated human neutrophils but not in activated T cells. Blood. 109, 219–27.
Nieminen J, Kuno A, Hirabayashi J, Sato S (2007) Visualization of galectin-3 oligomerization on the surface of neutrophils and endothelial cells using fluorescence resonance energy transfer. J Biol Chem. 282, 1374–83.
Lau KS, Partridge EA, Grigorian A, Silvescu CI, Reinhold VN, Demetriou M, Dennis JW (2007) Complex N-glycan number and degree of branching cooperate to regulate cell proliferation and differentiation. Cell. 129, 123–34.
Paz I, Sachse M, Dupont N, Mounier J, Cederfur C, Enninga J, Leffler H, Poirier F, Prevost MC, Lafont F, Sansonetti P (2009) Galectin-3, a marker for vacuole lysis by invasive pathogens. Cell Microbiol. 12, 530–44.
Dupont N, Lacas-Gervais S, Bertout J, Paz I, Freche B, Van Nhieu GT, van der Goot FG, Sansonetti PJ, Lafont F (2009) Shigella phagocytic vacuolar membrane remnants participate in the cellular response to pathogen invasion and are regulated by autophagy. Cell Host Microbe. 6, 137–49.
Thurston TL, Wandel MP, Muhlinen von N, Foeglein A, Randow F (2012) Galectin 8 targets damaged vesicles for autophagy to defend cells against bacterial invasion. Nature. 482, 414–8.
MacKinnon AC, Gibbons MA, Farnworth SL, Leffler H, Nilsson UJ, Delaine T, Simpson AJ, Forbes SJ, Hirani N, Gauldie J, Sethi T (2012) Regulation of transforming growth factor-β1-driven lung fibrosis by galectin-3. Am J Resp Crit Care Med. 185, 537–46.
Lin Y-H, Qiu D-C, Chang W-H, Yeh Y-Q, Jeng U-S, Liu F-T, Huang J-R (2017) The intrinsically disordered N-terminal domain of galectin-3 dynamically mediates multisite self-association of the protein through fuzzy interactions. J Biol Chem. 292, 17845–56.
Ludwig A-K, Michalak M, Xiao Q, Gilles U, Medrano FJ, Ma H, FitzGerald FG, Hasley WD, Melendez-Davila A, Liu M, Rahimi K, Kostina NY, Rodriguez-Emmenegger C, Möller M, Lindner I, Kaltner H, Cudic M, Reusch D, Kopitz J, Romero A, Oscarson S, Klein ML, Gabius H-J, Percec V (2019) Design-functionality relationships for adhesion/growth-regulatory galectins. Proc Natl Acad Sci U S A. 116, 2837–42.