Kunz C, Rudloff S, Baier W, Klein N, Strobel S (2000) Oligosaccharides in human milk: structural, functional, and metabolic aspects. Annu Rev Nutr, 20, 699-722.
McGuire MK, Meehan CL, McGuire MA, Williams JE, Foster J, Sellen DW, Kamau-Mbuthia EW, Kamundia EW, Mbugua S, Moore SE, Prentice AM, Kvist LJ, Otoo GE, Brooker SL, Price WJ, Shafii B, Placek C, Lackey KA, Robertson B, Manzano S, Ruíz L, Rodríguez JM, Pareja RG, Bode L (2017) What's normal? Oligosaccharide concentrations and profiles in milk produced by healthy women vary geographically. Am J Clin Nutr, 105, 1086-100.
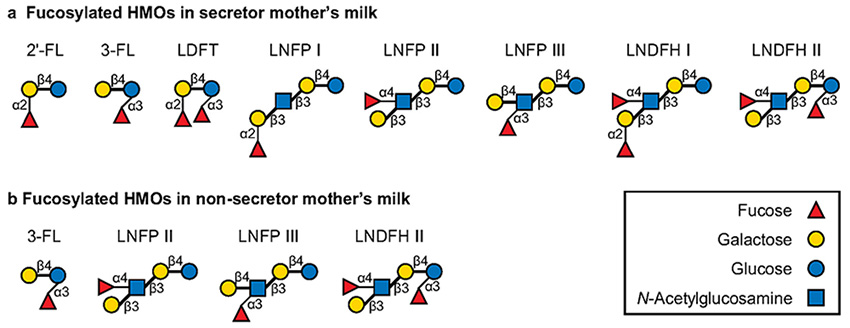
Urashima T, Asakuma S, Leo F, Fukuda K, Messer M, Oftedal OT (2012) The predominance of type I oligosaccharides is a feature specific to human breast milk. Adv Nutr, 3, 473S-82S.
Sakanaka M, Gotoh A, Yoshida K, Odamaki T, Koguchi H, Xiao JZ, Kitaoka M, Katayama T (2020) Varied pathways of infant gut-associated Bifidobacterium to assimilate human milk oligosaccharides: prevalence of the gene set and its correlation with bifidobacteria-rich microbiota formation. Nutrients, 12, 71.
Thomson P, Medina DA, Garrido D (2018) Human milk oligosaccharides and infant gut bifidobacteria: molecular strategies for their utilization. Food Microbiol, 75, 37-46.
Ojima MN, Yoshida K, Sakanaka M, Jiang L, Odamaki T, Katayama T (2022) Ecological and molecular perspectives on responders and non-responders to probiotics and prebiotics. Curr Opin Biotechnol, 73, 108-20.
Matsuki T, Yahagi K, Mori H, Matsumoto H, Hara T, Tajima S, Ogawa E, Kodama H, Yamamoto K, Yamada T, Matsumoto S, Kurokawa K (2016) A key genetic factor for fucosyllactose utilization affects infant gut microbiota development. Nat Commun, 7, 11939.
Lewis ZT, Totten SM, Smilowitz JT, Popovic M, Parker E, Lemay DG, Van Tassell ML, Miller MJ, Jin YS, German JB, Lebrilla CB, Mills DA (2015) Maternal fucosyltransferase 2 status affects the gut bifidobacterial communities of breastfed infants. Microbiome, 3, 13.
Berger B, Porta N, Foata F, Grathwohl D, Delley M, Moine D, Charpagne A, Siegwald L, Descombes P, Alliet P, Puccio G, Steenhout P, Mercenier A, Sprenger N (2020) Linking human milk oligosaccharides, infant fecal community types, and later risk to require antibiotics. mBio, 11, e03196-19.
Katayama T, Sakuma A, Kimura T, Makimura Y, Hiratake J, Sakata K, Yamanoi T, Kumagai H, Yamamoto K (2004) Molecular cloning and characterization of Bifidobacterium bifidum 1,2-α-ʟ-fucosidase (AfcA), a novel inverting glycosidase (glycoside hydrolase family 95). J Bacteriol, 186, 4885-93.
Ashida H, Miyake A, Kiyohara M, Wada J, Yoshida E, Kumagai H, Katayama T, Yamamoto K (2009) Two distinct α-ʟ-fucosidases from Bifidobacterium bifidum are essential for the utilization of fucosylated milk oligosaccharides and glycoconjugates. Glycobiology, 19, 1010-7.
Katoh T, Ojima MN, Sakanaka M, Ashida H, Gotoh A, Katayama T (2020) Enzymatic adaptation of Bifidobacterium bifidum to host glycans, viewed from glycoside hydrolyases and carbohydrate-binding modules. Microorganisms, 8, 481.
Gotoh A, Katoh T, Sakanaka M, Ling Y, Yamada C, Asakuma S, Urashima T, Tomabechi Y, Katayama-Ikegami A, Kurihara S, Yamamoto K, Harata G, He F, Hirose J, Kitaoka M, Okuda S, Katayama T (2018) Sharing of human milk oligosaccharides degradants within bifidobacterial communities in faecal cultures supplemented with Bifidobacterium bifidum. Sci Rep, 8, 13958.
Laursen MF, Sakanaka M, von Burg N, Mörbe U, Andersen D, Moll JM, Pekmez CT, Rivollier A, Michaelsen KF, Mølgaard C, Lind MV, Dragsted LO, Katayama T, Frandsen HL, Vinggaard AM, Bahl MI, Brix S, Agace W, Licht TR, Roager HM (2021) Bifidobacterium species associated with breastfeeding produce aromatic lactic acids in the infant gut. Nat Microbiol, 6, 1367-82.
Tsukuda N, Yahagi K, Hara T, Watanabe Y, Matsumoto H, Mori H, Higashi K, Tsuji H, Matsumoto S, Kurokawa K, Matsuki T (2021) Key bacterial taxa and metabolic pathways affecting gut short-chain fatty acid profiles in early life. ISME J, 15, 2574-90.
Vatanen T, Franzosa EA, Schwager R, Tripathi S, Arthur TD, Vehik K, Lernmark Å, Hagopian WA, Rewers MJ, She JX, Toppari J, Ziegler AG, Akolkar B, Krischer JP, Stewart CJ, Ajami NJ, Petrosino JF, Gevers D, Lähdesmäki H, Vlamakis H, Huttenhower C, Xavier RJ (2018) The human gut microbiome in early-onset type 1 diabetes from the TEDDY study. Nature, 562, 589-94.
Stewart CJ, Ajami NJ, O'Brien JL, Hutchinson DS, Smith DP, Wong MC, Ross MC, Lloyd RE, Doddapaneni H, Metcalf GA, Muzny D, Gibbs RA, Vatanen T, Huttenhower C, Xavier RJ, Rewers M, Hagopian W, Toppari J, Ziegler AG, She JX, Akolkar B, Lernmark A, Hyoty H, Vehik K, Krischer JP, Petrosino JF (2018) Temporal development of the gut microbiome in early childhood from the TEDDY study. Nature, 562, 583-88.
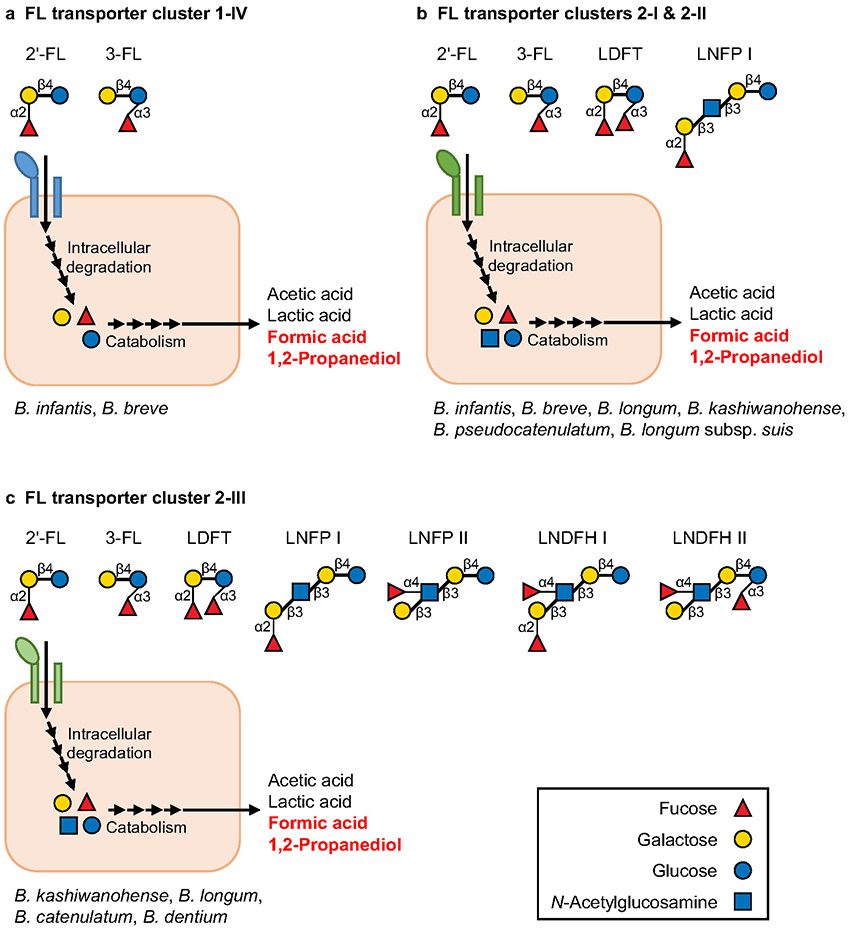
Sakanaka M, Hansen ME, Gotoh A, Katoh T, Yoshida K, Odamaki T, Yachi H, Sugiyama Y, Kurihara S, Hirose J, Urashima T, Xiao JZ, Kitaoka M, Fukiya S, Yokota A, Lo Leggio L, Abou Hachem M, Katayama T (2019) Evolutionary adaptation in fucosyllactose uptake systems supports bifidobacteria-infant symbiosis. Sci Adv, 5, eaaw7696.
Ojima MN, Asao Y, Nakajima A, Katoh T, Kitaoka M, Gotoh A, Hirose J, Urashima T, Fukiya S, Yokota A, Abou Hachem M, Sakanaka M, Katayama T (2022) Diversification of a fucosyllactose transporter within the genus Bifidobacterium. Appl Environ Microbiol, 88: e01437-21.
Garrido D, Ruiz-Moyano S, Kirmiz N, Davis JC, Totten SM, Lemay DG, Ugalde JA, German JB, Lebrilla CB, Mills DA (2016) A novel gene cluster allows preferential utilization of fucosylated milk oligosaccharides in Bifidobacterium longum subsp. longum SC596. Sci Rep, 6, 35045.
James K, Bottacini F, Contreras JIS, Vigoureux M, Egan M, O'Connell-Motherway M, Holmes E, van Sinderen D (2019) Metabolism of the predominant human milk oligosaccharide fucosyllactose by an infant gut commensal. Sci Rep, 9, 15427.
Shani G, Hoeflinger JL, Heiss BE, Masarweh CF, Larke JA, Jensen NM, Wickramasinghe S, Davis JC, Goonatilleke E, El-Hawiet A, Nguyen L, Klassen JS, Slupsky CM, Lebrilla CB, Mills DA (2022) Fucosylated human milk oligosaccharide foraging within the species Bifidobacterium pseudocatenulatum is driven by glycosyl hydrolase content and specificity. Appl Environ Microbiol, 88: e01707-21.
Bunesova V, Lacroix C, Schwab C (2016) Fucosyllactose and L-fucose utilization of infant Bifidobacterium longum and Bifidobacterium kashiwanohense. BMC Microbiol, 16, 248.
Dedon LR, Özcan E, Rani A, Sela DA (2020) Bifidobacterium infantis metabolizes 2ʹfucosyllactose-derived and free fucose through a common catabolic pathway resulting in 1,2-propanediol secretion. Front Nutr, 7, 583397.
Zabel B, Yde CC, Roos P, Marcussen J, Jensen HM, Salli K, Hirvonen J, Ouwehand AC, Morovic W (2019) Novel genes and metabolite trends in Bifidobacterium longum subsp. infantis Bi-26 metabolism of human milk oligosaccharide 2ʹ-fucosyllactose. Sci Rep, 9, 7983.
Cheng CC, Duar RM, Lin X, Perez-Munoz ME, Tollenaar S, Oh JH, van Pijkeren JP, Li F, van Sinderen D, Gänzle MG, Walter J (2020) Ecological importance of cross-feeding of the intermediate metabolite 1,2-propanediol between bacterial gut symbionts. Appl Environ Microbiol, 86, e00190-20.
Egan M, O'Connell-Motherway M, Kilcoyne M, Kane M, Joshi L, Ventura M, van Sinderen D (2014) Cross-feeding by Bifidobacterium breve UCC2003 during co-cultivation with Bifidobacterium bifidum PRL2010 in a mucin-based medium. BMC Microbiol, 14, 282.
Higgins MA, Ryan KS (2021) Generating a fucose permease deletion mutant in Bifidobacterium longum subspecies infantis ATCC 15697. Anaerobe, 68, 102320.
Heiss BE, Ehrlich AM, Maldonado-Gomez MX, Taft DH, Larke JA, Goodson ML, Slupsky CM, Tancredi DJ, Raybould HE, Mills DA (2021) Bifidobacterium catabolism of human milk oligosaccharides overrides endogenous competitive exclusion driving colonization and protection. Gut Microbes, 13, 1986666.
Duar RM, Kyle D, Casaburi G (2020) Colonization resistance in the infant gut: the role of B. infantis in reducing pH and preventing pathogen growth. High Throughput, 9, 7.
Schwab C, Ruscheweyh HJ, Bunesova V, Pham VT, Beerenwinkel N, Lacroix C (2017) Trophic interactions of infant bifidobacteria and Eubacterium hallii during ʟ-fucose and fucosyllactose degradation. Front Microbiol, 8, 95.
Li M, Bai Y, Zhou J, Huang W, Yan J, Tao J, Fan Q, Liu Y, Mei D, Yan Q, Yuan J, Malard P, Wang Z, Gu J, Tanigchi N, Li W (2019) Core fucosylation of maternal milk N-glycan evokes B cell activation by selectively promoting the ʟ-fucose metabolism of gut Bifidobacterium spp. and Lactobacillus spp. mBio, 10, e00128-19.
Yu ZT, Chen C, Newburg DS (2013) Utilization of major fucosylated and sialylated human milk oligosaccharides by isolated human gut microbes. Glycobiology, 23, 1281-92.
Kostopoulos I, Elzinga J, Ottman N, Klievink JT, Blijenberg B, Aalvink S, Boeren S, Mank M, Knol J, de Vos WM, Belzer C (2020) Akkermansia muciniphila uses human milk oligosaccharides to thrive in the early life conditions in vitro. Sci Rep, 10, 14330.
Pichler MJ, Yamada C, Shuoker B, Alvarez-Silva C, Gotoh A, Leth ML, Schoof E, Katoh T, Sakanaka M, Katayama T, Jin C, Karlsson NG, Arumugam M, Fushinobu S, Abou Hachem M (2020) Butyrate producing colonic Clostridiales metabolise human milk oligosaccharides and cross feed on mucin via conserved pathways. Nat Commun, 11, 3285.