Blix G. (1936) Über die Kohlenhydratgruppen des Submaxillarismucins. 240:43–54.
Kooner AS, Yu H, Chen X. (2019) Synthesis of N-Glycolylneuraminic Acid (Neu5Gc) and Its Glycosides. Front. Immunol. 10:2004.
van Karnebeek CDM, Bonafé L, Wen X-Y, Tarailo-Graovac M, Balzano S, Royer-Bertrand B, Ashikov A, Garavelli L, Mammi I, Turolla L, Breen C, Donnai D, Cormier-Daire V, Heron D, Nishimura G, Uchikawa S, Campos-Xavier B, Rossi A, Hennet T, Brand-Arzamendi K, Rozmus J, Harshman K, Stevenson BJ, Girardi E, Superti-Furga G, Dewan T, Collingridge A, Halparin J, Ross CJ, Van Allen MI, Rossi A, Engelke UF, Kluijtmans LAJ, van der Heeft E, Renkema H, de Brouwer A, Huijben K, Zijlstra F, Heise T, Boltje T, Wasserman WW, Rivolta C, Unger S, Lefeber DJ, Wevers RA, Superti-Furga A. (2016) NANS-mediated synthesis of sialic acid is required for brain and skeletal development. Nat. Genet. 48:777–784.
Varki A. (2008) Sialic acids in human health and disease. Trends Mol. Med. 14:351–360.
Lewis AL, Lewis WG. (2012) Host sialoglycans and bacterial sialidases: A mucosal perspective. Cell. Microbiol. 14:1174–1182.
Urashima T. (2021) Human milk oligosaccharides. Glycoforum 24:A14J.
Wang B. (2009). Sialic acid is an essential nutrient for brain development and cognition. Annu. Rev. Nutr. 29:177–222.
Zhang X, Liu Y, Liu L, Li J, Du G, Chen J. (2019) Microbial production of sialic acid and sialylated human milk oligosaccharides: Advances and perspectives. Biotechnol. Adv. 37:787–800.
Martín-Sosa S, Martín M-J, García-Pardo LA, Hueso P. (2004) Distribution of sialic acids in the milk of Spanish mothers of full term infants during lactation. J. Pediatr. Gastroenterol. Nutr. 39:499–503.
Brockhausen I, Schachter H, Stanley P. O-GalNAc Glycans. In: Varki A, Cummings RD, Esko JD, Freeze HH, Stanley P, Bertozzi CR, Hart GW, Etzler ME. (2009) Essentials of Glycobiology 2nd edition. Cold Spring Harbor Laboratory Press, Cold Spring Harbor (NY).
Theodoratou E, Campbell H, Ventham NT, Kolarich D, Pučić-Baković M, Zoldoš V, Fernandes D, Pemberton IK, Rudan I, Kennedy NA, Wuhrer M, Nimmo E, Annese V, McGovern DPB, Satsangi J, Lauc G. (2014) The role of glycosylation in IBD. Nat. Rev. Gastroenterol. Hepatol. 11:588–600.
Hopkins AP, Hawkhead JA, Thomas GH. (2013) Transport and catabolism of the sialic acids N-glycolylneuraminic acid and 3-keto-3-deoxy-d-glycero-d-galactonononic acid by Escherichia coli K-12. FEMS Microbiol. Lett. 347:14–22.
Severi E, Rudden M, Bell A, Palmer T, Juge N, Thomas GH. (2021) Multiple evolutionary origins reflect the importance of sialic acid transporters in the colonization potential of bacterial pathogens and commensals. Microb. Genomics 7:000614.
Almagro-Moreno S, Boyd EF. (2009) Insights into the evolution of sialic acid catabolism among bacteria. BMC Evol. Biol. 9:118.
Ravcheev DA, Thiele I. (2017) Comparative genomic analysis of the human gut microbiome reveals a broad distribution of metabolic pathways for the degradation of host-synthetized mucin glycans and utilization of mucin-derived monosaccharides. Front. Genet. 8:111.
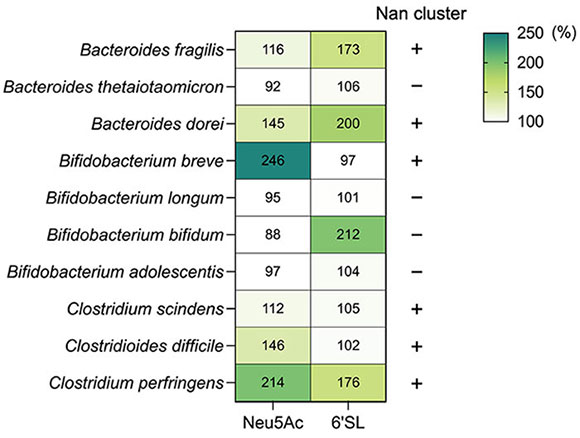
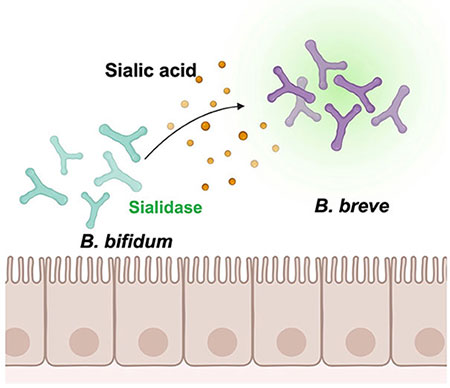
Nishiyama K, Nagai A, Uribayashi K, Yamamoto Y, Mukai T, Okada N. (2018) Two extracellular sialidases from Bifidobacterium bifidum promote the degradation of sialyl-oligosaccharides and support the growth of Bifidobacterium breve. Anaerobe 52:22–28.
Duncan PI, Raymond F, Fuerholz A, Sprenger N. (2009) Sialic acid utilisation and synthesis in the neonatal rat revisited. PLoS One 4:e8241.
Akazawa H, Tsujikawa Y, Fukuda I, Suzuki Y, Choi M, Katayama T, Mukai T, Osawa R. (2021) Isolation and identification of milk oligosaccharide-degrading bacteria from the intestinal contents of suckling rats. Biosci. Microbiota Food Health 40:27–32.
Matsui S, Akazawa H, Tsujikawa Y, Fukuda I, Suzuki Y, Yamamoto Y, Mukai T, Shirai Y, Osawa R. (2021) Milk oligosaccharide-mediated cross-feeding between Enterococcus gallinarum and lactobacilli in the gut microbiota of infant rats. Biosci. Microbiota Food Health 40:204–211.
Lynch JB, Sonnenburg JL. (2012) Prioritization of a plant polysaccharide over a mucus carbohydrate is enforced by a Bacteroides hybrid two-component system. Mol. Microbiol. 85:478–491.
Nishiyama K, Yokoi T, Sugiyama M, Osawa R, Mukai T, Okada N. (2021) Roles of the cell surface architecture of Bacteroides and Bifidobacterium in the gut colonization. Front. Microbiol. 12:754819.
Ng KM, Ferreyra JA, Higginbottom SK, Lynch JB, Kashyap PC, Gopinath S, Naidu N, Choudhury B, Weimer BC, Monack DM, Sonnenburg JL. (2013) Microbiota-liberated host sugars facilitate post-antibiotic expansion of enteric pathogens. Nature 502:96–99.
Charbonneau MR, O’Donnell D, Blanton LV, Totten SM, Davis JCC, Barratt MJ, Cheng J, Guruge J, Talcott M, Bain JR, Muehlbauer MJ, Ilkayeva O, Wu C, Struckmeyer T, Barile D, Mangani C, Jorgensen J, Fan YM, Maleta K, Dewey KG, Ashorn P, Newgard CB, Lebrilla C, Mills DA, Gordon JI. (2016) Sialylated milk oligosaccharides promote microbiota-dependent growth in models of infant undernutrition. Cell 164:859–871.
Cowardin CA, Ahern PP, Kung VL, Hibberd MC, Cheng J, Guruge JL, Sundaresan V, Head RD, Barile D, Mills DA, Barratt MJ, Huq S, Ahmed T, Gordon JI. (2019) Mechanisms by which sialylated milk oligosaccharides impact bone biology in a gnotobiotic mouse model of infant undernutrition. Proc. Natl. Acad. Sci. U. S. A. 116:11988–11996.
Perdijk O, van Baarlen P, Fernandez-Gutierrez MM, van den Brink E, Schuren FHJ, Brugman S, Savelkoul HFJ, Kleerebezem M, van Neerven RJJ. (2019) Sialyllactose and galactooligosaccharides promote epithelial barrier functioning and distinctly modulate microbiota composition and short chain fatty acid production In vitro. Front. Immunol. 10:94.
Park S-Y, Rao C, Coyte KZ, Kuziel GA, Zhang Y, Huang W, Franzosa EA, Weng J-K, Huttenhower C, Rakoff-Nahoum S. (2022) Strain-level fitness in the gut microbiome is an emergent property of glycans and a single metabolite. Cell 185:513-529.e21.
Bergfeld AK, Pearce OMT, Diaz SL, Pham T, Varki A. (2012) Metabolism of vertebrate amino sugars with N-glycolyl groups: elucidating the intracellular fate of the non-human sialic acid N-glycolylneuraminic acid. J. Biol. Chem. 287:28865–28881.
Chen Y, Pan L, Liu N, Troy FA, Wang B. (2014) LC-MS/MS quantification of N-acetylneuraminic acid, N-glycolylneuraminic acid and ketodeoxynonulosonic acid levels in the urine and potential relationship with dietary sialic acid intake and disease in 3- to 5-year-old children. Br. J. Nutr. 111:332–341.
Samraj AN, Pearce OMT, Läubli H, Crittenden AN, Bergfeld AK, Banda K, Gregg CJ, Bingman AE, Secrest P, Diaz SL, Varki NM, Varki A. (2015) A red meat-derived glycan promotes inflammation and cancer progression. Proc. Natl. Acad. Sci. U. S. A. 112:542–547.
Zaramela LS, Martino C, Alisson-Silva F, Rees SD, Diaz SL, Chuzel L, Ganatra MB, Taron CH, Secrest P, Zuñiga C, Huang J, Siegel D, Chang G, Varki A, Zengler K. (2019) Gut bacteria responding to dietary change encode sialidases that exhibit preference for red meat-associated carbohydrates. Nat. Microbiol. 4:2082–2089.