上記のような背景から、人乳にはビフィズス菌を増殖させる成分「ビフィズス因子」が含まれると考えられるようになり、1910年頃から関連する研究が精力的に行われた。1953年にGyörgyらは、Bifidobacterium bifidum var. Pennsilvanicus株(当時の分類ではLactobacillus bifidus)の増殖促進能を指標に人乳からビフィズス因子の単離を行い、それはN-アセチルグルコサミン(GlcNAc)、フコース(Fuc)、ガラクトース(Gal)、およびグルコース(Glc)からなるラクトース以外のオリゴ糖であると報告した5。この報告以降、母乳に含まれる三糖以上のオリゴ糖混合物であるヒトミルクオリゴ糖がビフィズス因子であると考えられるようになった。
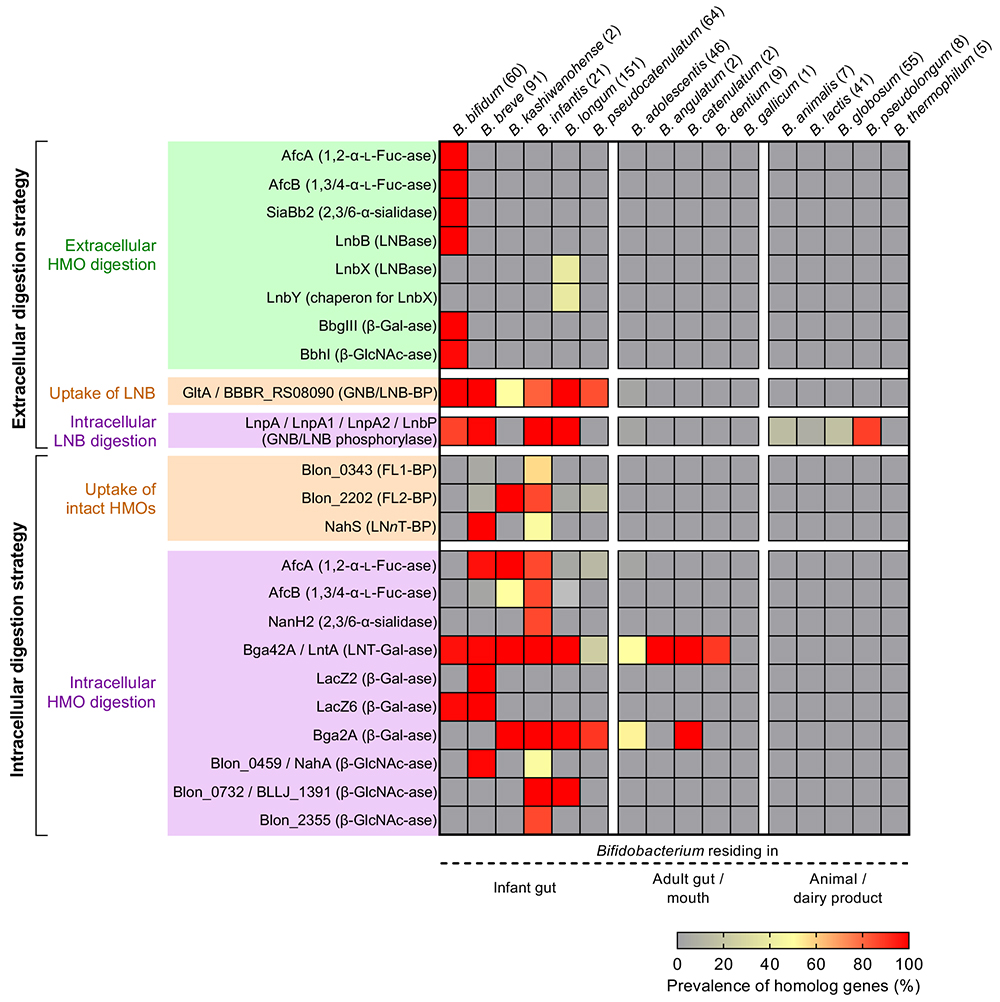
このラクト-N-ビオースI仮説を契機としてHMO分解に関わる酵素群が次々に発見され、現在までにHMO分解に必要なビフィズス菌の全てのグリコシダーゼ群と一部のトランスポーターが同定されている(4 . HMO資化に関わるビフィズス菌の酵素を参照)。なお、同時期にDavid A. MillsらのグループがB. infantisに高いHMO資化性を発見しており、彼らはその後、基準株ATCC 15697株(= JCM 1222株)のゲノム解析を行うことでHMO分解の分子基盤解明に取り組んだ12。
4. HMO資化に関わるビフィズス菌の酵素
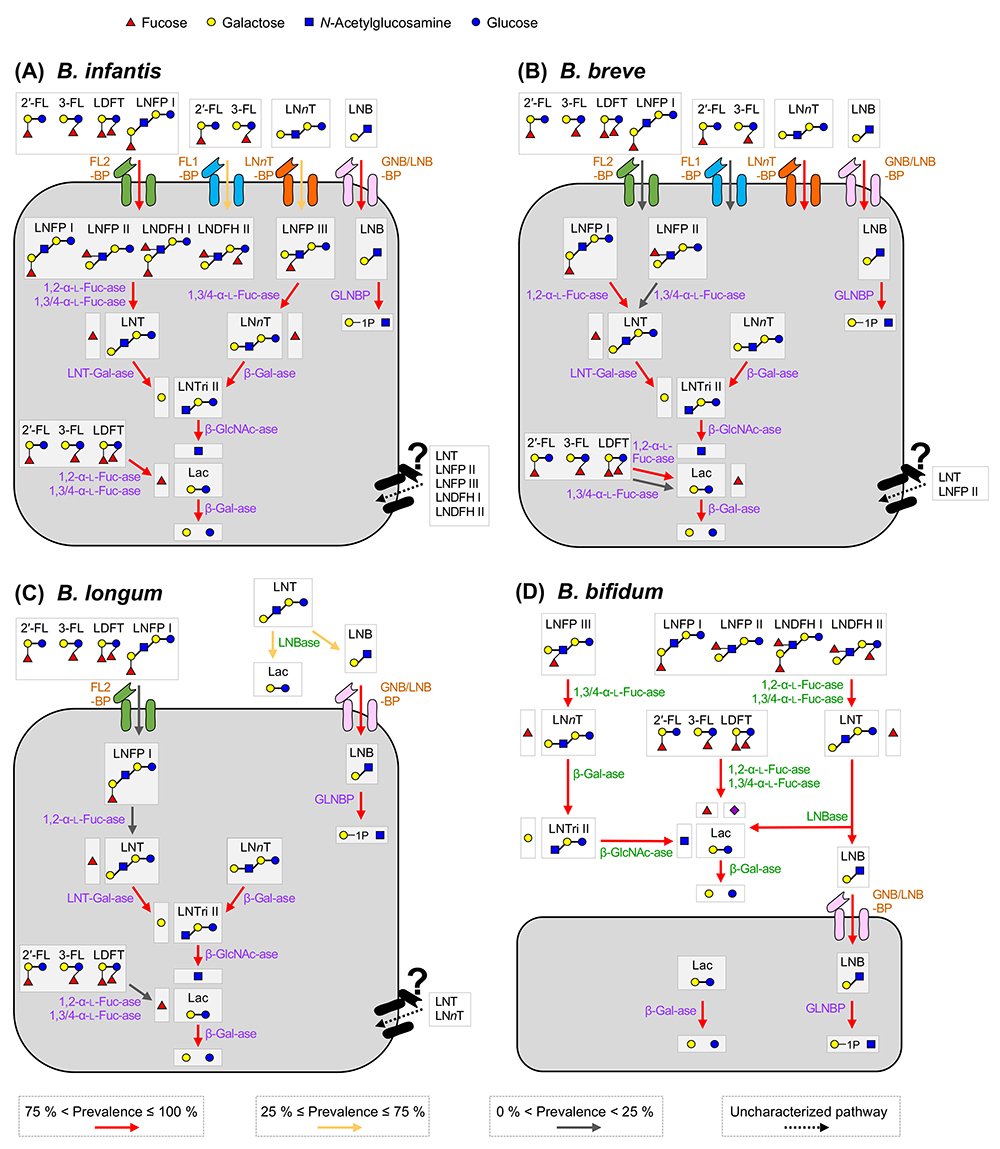
代表的な乳児型ビフィズス菌B. breve, B. infantis, B. longum, およびB. bifidumのHMO分解経路の概要を図 1に示す。酵素の詳細について以下に説明するが、ビフィズス菌のHMO資化経路は大きく分けて菌体外で単糖・2糖にまで分解するタイプと3糖以上の構造を直接取込むタイプに分けられる。
図 1. 主要な乳児型ビフィズス菌種におけるHMO資化経路10種類の代表的な中性HMOのB. infantis (A), B. breve (B), B. longum (C), およびB. bifidum (D)における資化経路を示す。該当する酵素の種内保有率を異なる色の矢印で示した(保有率については図 3を参照)。赤色矢印は75 %以上の、黄色は25−75 %の、灰色は25 %以下を表す。未同定の経路については点線で示した。HMOを菌体表層で分解して利用するタイプにかかわる細胞壁アンカー型酵素を薄緑色で、トランスポーターを薄茶色で、HMOを直接取込み利用するタイプにかかわる菌体内酵素は薄紫色で示した。細胞外で生じた単糖・2糖は他の腸内細菌、特にビフィズス菌内にクロスフィードされる。
本図は、阪中らの論文29を一部改変して作成した。
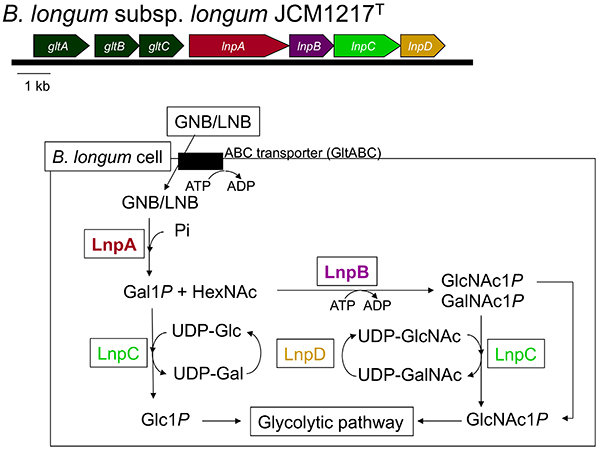
まとめると、GNB/LNB経路は菌体外で生成したGNB/LNBを菌体内に取り込んで分子全体を解糖系に送り込むための経路である。GNB/LNB経路は当時調べた限りの乳児型ビフィズス菌B. breve, B. infantis, B. longum, およびB. bifidumの全ての株が保持していたが、成人糞便から単離されるB. adolescentis, B. catenulatum,およびBifidobacterium animalis subsp. lactisは保持していなかった11。
Homma N (1994) History of the research on bifidobacteria. in Research on Bifidobacteria (Mitsuoka T ed), Japan Bifidus Foundation, pp. 1-22 (in Japanese).
Sakanaka M, Hansen ME, Gotoh A, Katoh T, Yoshida K, Odamaki T, Yachi H, Sugiyama Y, Kurihara S, Hirose J, Urashima T, Xiao JZ, Kitaoka M, Fukiya S, Yokota A, Lo Leggio L, Hachem MA, and Katayama T (2019) Evolutionary adaptation in fucosyllactose uptake systems supports bifidobacteria-infant symbiosis. Sci Adv, 5, eaaw7696.
Berger B, Porta N, Foata F, Grathwohl D, Delley M, Moine D, Charpagne A, Siegwald L, Descombes P, Alliet P, Puccio G, Steenhout P, Mercenier A, Sprenger N (2020) Linking human milk oligosaccharides, infant fecal community types, and later risk to require antibiotics. mBio, 11, e03196-19.
Gibson GR, Hutkins R, Sanders ME, Prescott SL, Reimer RA, Salminen SJ, Scott K, Stanton C, Swanson KS, Cani PD, Verbeke K, Reid G (2017) Expert consensus document: The International Scientific Association for Probiotics and Prebiotics (ISAPP) consensus statement on the definition and scope of prebiotics. Nature Rev Gastroenterol Hepatol, 14, 491-502.
György P, Kuhn R, Rose CS, Zilliken F (1954) Bifidus factor. II. Its occurrence in milk from different species and in other natural products. Arch Biochem Biophys, 48, 202-8.
Urashima T, Asakuma S, Leo F, Fukuda K, Messer M, Oftedal OT (2012) The predominance of type I oligosaccharides is a feature specific to human breast milk. Adv Nutr, 3, 473S-482S.
Derensy-Dron D, Krzewinski F, Brassart C, Bouquelet S (1999) β-1,3-Galactosyl-N-acetylhexosamine phosphorylase from Bifidobacterium bifidum DSM 20082: characterization, partial purification and relation to mucin degradation. Biotechnol Appl Biochem, 29, 3-10.
Kitaoka M, Tian J, Nishimoto M (2005) Novel putative galactose operon involving lacto-N-biose phosphorylase in Bifidobacterium longum. Appl Environ Microbiol, 71, 3158-62.
Nishimoto M, Kitaoka M (2007) Identification of the putative proton donor residue of lacto-N-biose phosphorylase (EC 2.4.1.211). Biosci Biotechnol Biochem, 71, 1587-91.
Xiao J, Takahashi S, Nishimoto M, Odamaki T, Yaeshima T, Iwatsuki K, Kitaoka M (2010) Distribution of in vitro fermentation ability of lacto-N-biose I, the major building block of human milk oligosaccharides, in bifidobacterial strains. Appl Environ Microbiol, 76, 54-9.
Sela DA, Chapman J, Adeuya A, Kim JH, Chen F, Whitehead TR, Lapidus A, Rokhsar DS, Lebrilla CB, German JB, Price NP, Richardson PM, Mills DA (2008) The genome sequence of Bifidobacterium longum subsp. infantis reveals adaptations for milk utilization within the infant microbiome. Proc Natl Acad Sci USA, 105, 18964-9.
Suzuki R, Wada J, Katayama T, Fushinobu S, Wakagi T, Shoun H, Sugimoto H, Tanaka A, Kumagai H, Ashida H, Kitaoka M, Yamamoto K (2008) Structural and thermodynamic analyses of solute-binding protein from Bifidobacterium longum specific for core 1 disaccharide and lacto-N-biose I. J Biol Chem, 283, 13165-73.
Nishimoto M, Kitaoka M (2007) Identification of N-acetylhexosamine 1-kinase in the complete lacto-N-biose I/galacto-N-biose metabolic pathway in Bifidobacterium longum. Appl Environ Microbiol, 73, 6444-9.
Yamada C, Gotoh A, Sakanaka M, Hattie M, Stubbs KA, Katayama-Ikegami A, Hirose J, Kurihara S, Arakawa T, Kitaoka M, Okuda S, Katayama T, Fushinobu S (2017) Molecular insight into evolution of symbiosis between breast-fed infants and a member of the human gut microbiome Bifidobacterium longum. Cell Chem Biol, 24, 515-524.
Sano M, Hayakawa K, Kato I (1993) Purification and characterization of an enzyme releasing lacto-N-biose from oligosaccharides with type 1 chain. J Biol Chem, 268, 18560-6.
Wada J, Ando T, Kiyohara M, Ashida H, Kitaoka M, Yamaguchi M, Kumagai H, Katayama T, Yamamoto K (2008) Bifidobacterium bifidum lacto-N-biosidase, a critical enzyme for the degradation of human milk oligosaccharides with a type-1 structure. Appl Environ Microbiol, 74, 3996-4004.
Sakurama H, Kiyohara M, Wada J, Honda Y, Yamaguchi M, Fukiya S, Yokota A, Ashida H, Kumagai H, Kitaoka M, Yamamoto K, Katayama T (2013) Lacto-N-biosidase encoded by a novel gene of Bifidobacterium longum subspecies longum shows unique substrate specificity and requires a designated chaperone for its active expression. J Biol Chem, 288, 25194-206.
Katayama T, Sakuma A, Kimura T, Makimura Y, Hiratake J, Sakata K, Yamanoi T, Kumagai H, Yamamoto K (2004) Molecular cloning and characterization of Bifidobacterium bifidum 1,2-α-L-fucosidase (AfcA), a novel inverting glycosidase (glycoside hydrolase family 95). J Bacteriol, 186, 4885-93.
Ashida H, Miyake A, Kiyohara M, Wada J, Yoshida E, Kumagai H, Katayama T, Yamamoto K (2009) Glycobiology, 19, 1010-17.
Nishiyama K, Yamamoto Y, Sugiyama M, Takaki T, Urashima T, Fukiya S, Yokota A, Oada N, Mukai T (2017) Bifidobacterium bifidum extracellular sialidase enhances adhesion to the mucosal surface and supports carbohydrate assimilation. mBio, 8, e00928-17.
Yoshida E, Sakurama H, Kiyohara M, Nakajima M, Kitaoka M, Ashida H, Hirose J, Katayama T, Yamamoto K, Kumagai H (2012) Bifidobacterium longum subsp. infantis uses two different β-galactosidases for selectively degrading type-1 and type-2 human milk oligosaccharides. Glycobiology, 22, 361-8.
Garrido D, Ruiz-Moyano S, Mills DA (2012) Release and utilization of N-acetyl-D-glucosamine from human milk oligosaccharides by Bifidobacterium longum subsp. infantis. Anaerobe, 18, 430–5.
Honda Y, Nishimoto M, Katayama T, Kitaoka M (2013) Characterization of the cytosolic β-N-acetylglucosaminidase from Bifidobacterium longum subsp. longum. J Appl Glycosci, 60, 141-6.
Asakuma S, Hatakeyama E, Urashima T, Yoshida E, Katayama T, Yamamoto K, Kumagai H, Ashida H, Hirose J, Kitaoka M (2011) Physiology of consumption of human milk oligosaccharides by infant gut-associated bifidobacteria. J Biol Chem, 286, 34583-92.
Gotoh A, Katoh T, Sakanaka M, Ling Y, Yamada C, Asakuma S, Urashima T, Tomabechi Y, Katayama-Ikegami A, Kurihara S, Yamamoto K, Harata G, He F, Hirose J, Kitaoka M, Okuda S, Katayama T (2018) Sharing of human milk oligosaccharides degradants within bifidobacterial communities in faecal cultures supplemented with Bifidobacterium bifidum. Sci Rep, 8, 13958.
Tannock GW, Lawley B, Munro K, Pathmanathan SG, Zhou SJ, Makrides M, Gibson RA, Sullivan T, Prosser CG, Lowry D, and Hodgkinsong AJ (2013) Comparison of the compositions of the stool microbiotas of infants fed goat milk Formula, cow milk-based formula, or breast milk. Appl Environ Microbiol, 79, 3040–8.
Sakanaka M, Gotoh A, Yoshida K, Odamaki T, Koguchi H, Xiao JZ, Kitaoka M, Katayama T (2020) Varied pathways of infant gut-associated Bifidobacterium to assimilate human milk oligosaccharides: prevalence of the gene set and its correlation with bifidobacteria-rich microbiota formation. Nutrients, 12, 7.
Marcobal A, Barboza M, Sonnenburg ED, Pudlo N, Martens EC, Desai P, Lebrilla CB, Weimer BC, Mills DA, German JB, Sonnenburg JL (2011) Bacteroides in the infant gut consume milk oligosaccharides via mucus-utilization pathways. Cell Host Microbe, 10, 507–14.
Kostopoulos I, Elzinga J, Ottman N, Klievink JT, Blijenberg B, Aalvink S, Boeren S, Mank M, Knol J, de Vos WM, Belzer C (2020) Akkermansia muciniphila uses human milk oligosaccharides to thrive in the early life conditions in vitro. Sci Rep, 10, 14330.
Pichler MJ, Yamada C, Shuoker B, Alvarez-Silva C, Gotoh A, Leth ML, Schoof E, Katoh T, Sakanaka M, Katayama T, Jin C, Karlsson NG, Arumugam M, Fushinobu S, Hachem MA (2020) Butyrate producing colonic Clostridiales metabolise human milk oligosaccharides and cross feed on mucin via conserved pathways. Nat Commun, 11, 3285.
Glick MC, Sall T, Zilliken F, Mudd S (1960) Morphological changes of Lactobacillus bifidus var. Pennsylvanicus produced by a cell-wall precursor. Biochim Biophys Acta, 37, 361-3.
Ojima MN, Yoshida K, Sakanaka M, Jiang L, Odamaki T, Katayama T (2022) Ecological and molecular perspectives on responders and non-responders to probiotics and prebiotics. Curr Opin Biotechnol, 73, 108-20.