

|
|
Immunity and the Sugar chain: in the web site "Glycoword" |
|||||||||||||||||||||||
 |
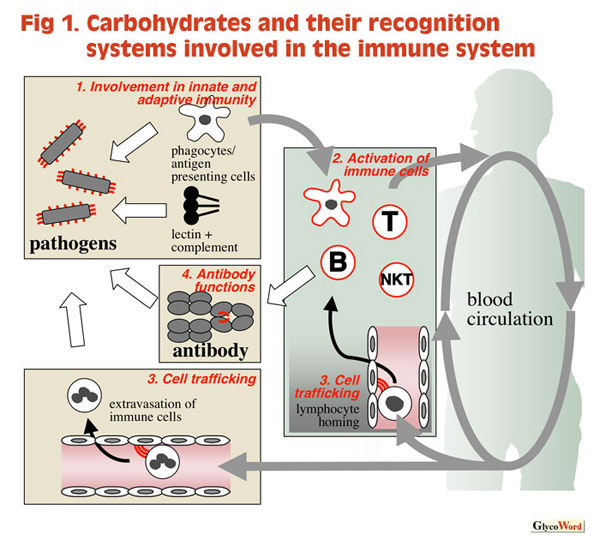
The immune system protects the body from infections and diseases by innate and adaptive immunity. Typical features of the immune system are its ability to recognize a variety of foreign substances and self-nonself discrimination. Other unique features involve many kinds of immune cells originating in bone marrow with their varied differentiation and activation stages, the traffic of immune cells throughout the body, as well as a number of positive and negative regulatory systems that facilitate fine tuning of the immune response. The web site "Glycoword" selects typical and recent topics on carbohydrates and recognition molecules that are involved in the immune system. This article summarizes the contents of the web site. |
||||||||||||||||||||||
| 1. Involvement of carbohydrate recognition in innate immunity A variety of carbohydrates and their recognition molecules are involved in the recognition of pathogens and non-self materials, reflecting long evolutionary processes of struggle between hosts and pathogens. Lipopolysaccharides, the principal outer membrane component of Gram-negative bacteria, is a kind of glycoconjugate that are recognized by pattern-recognition receptors including Toll-like receptors (Hashimoto, “LPS and Toll-like Receptor”). The host disposes of invading pathogens by use of humoral and cell-mediated protective mechanisms, both of which are triggered by lectin binding. The complement pathway is a humoral immune system that is triggered by the binding of collectins and mannose-binding proteins to pathogens, similar to antibody-mediated binding (Kawasaki “Collectins and Their Roles in Host Defense,” Matsushita “Mannose-binding Lectin and the Complement System,” Wakamiya “Collectins and Viral Infections”). Macrophages and dendritic cells express a set of membrane-bound C-type lectins. The lectin-mediated recognition of pathogens is greatly involved both in innate immunity for phagocytosis and in triggering adaptive immunity for efficient antigen uptake and presentation (Higashi “C-type Lectins Expressed on Macrophages and Dendritic Cells”). Cancer immunotherapy is an example of the applied manipulation of the acquired immunity system. MUC-1 has been studied extensively as a target molecule for the therapy (Nakada “Mucin and Immune Response”). |
|||||||||||||||||||||||
|
2. Glycoconjugates and carbohydrate recognition molecules that are involved in activation of immune cells
Each immune cell subset expresses different structures of glycoconjugates and their recognition molecules, which are often utilized as cell-specific and differentiation markers. Furthermore, some of them are actually involved in cell-type specific signal transduction (reference #1). Extracellular lectins transduce or modulate signal transduction. Galectin-3, originally identified as a galactoside-recognition molecule, modulates T cell receptor-dependent lymphocyte stimulation via binding to cell surface carbohydrate and subsequent forming of a lattice structure on the surface. Galectins, as soluble mediators, modulate many other biological functions such as migration, adhesion, and cytokine production (Sato, “Regulation of Immune Responses by Galectins”). It is also known that cytokines often possess carbohydrate binding activities that influence efficient signal transduction (reference #2). Glycoconjugates also participate in signal transduction. NKT cells are stimulated by a unique glycolipid galactosylceramide that are presented with CD1 molecules (Kawano and Taniguchi “V alpha 14 NKT Cells - A Novel Lymphocyte Specific for Glycolipid Antigens”). As an example of negative regulation, soluble mucin molecules induce secretion of prostaglandin E2 from macrophages and subsequently suppress immune function (Nakada “Mucin and Immune Response”). Siglecs are a family of immunoglobulin superfamily lectins that recognize glycans containing sialic acids. Each individual is expressed in a restricted population of immune cells. Their functions mostly remain unclear, although binding of CD22 on the B lymphocytes with its carbohydrate ligand is involved in the transduction of negative signal for the lymphocyte activation. (Angata “Siglecs (sialic acid-binding immunoglobulin superfamily lectins),” Takematsu “CD22(Siglec-2) and regulation of its glycan ligand expression”). |
|||||||||||||||||||||||
 |
|||||||||||||||||||||||
|
3. Carbohydrates and their recognition molecules in cell trafficking
Potential antigen presenting cells are located in the peripheral tissues for survey of antigen entry, whereas lymphocytes circulate throughout the body. In the phase of antigen presentation, both must move and meet in the same lymphoid organs such as lymph nodes to achieve efficient contact. In the effector phase, immune cells move and accumulate at the site of inflammation. Lectin-carbohydrate interaction partly regulates such complicated cell trafficking. Structure and functional roles of selectins and their carbohydrate ligands have been studied extensively, contributing much to the field of glycobiology and immunology (Imai “Cell Adhesion Molecules: Selectins,” Kannagi “Carbohydrate Determinants Involved in Inflammation and Homing of Lymphocytes,” Hiraoka “Glycosyltransferases, Sulfotransferases and Biosynthesis of L-Selectin Ligands”). Chemokines are soluble mediators that induce migration and accumulation of immune cells toward lymphoid organs and peripheral tissues. Directional migration of the immune cells can be achieved by site-specific distribution of the molecules that are often presented by proteoglycans that bind to them (Tanaka “Heparan Sulfate Proteoglycan in the Context of Cytokines and Adhesion Molecules,” reference #3). |
|||||||||||||||||||||||
|
4. Roles of carbohydrates in antibody functions
Antibodies are the principal effector molecules in humoral immunity. However, a few of them cause autoimmune diseases and allergy because antibodies to a pathogen may attack a mimetic glycan structure expressed in the body or on other foreign materials (Kusunoki “Immune-mediated Neuropathy and Antibody against Glycoconjugates,” reference #4). Altered carbohydrate structure in immunoglobulin is suspected to change the biochemical properties of molecules and to potentially cause some diseases, for example, altered N-glycan structure in the IgG constant region for rheumatoid arthritis (Endo “Rheumatoid Arthritis and Abnormality of Human Serum IgG N-Glycans”), and O-glycan in the hinge region of IgA for nephropathy (references #5, #6). On the other hand, artificial modification of N-glycan on IgG successfully elevates the activity of antibody-dependent cellular cytotoxicity. This is important for the improvement of antibody drugs (Shitara “Enhancement of ADCC of Antibodies by Glycoengineering”). Recent research efforts revealed that carbohydrates and their recognition systems have been finely incorporated in the present immune system as a result of long evolutionary processes. Hopefully, profound comprehension of the function of carbohydrates in the immune system will help generate new therapies for a number of immune diseases. |
|||||||||||||||||||||||
| Nobuaki Higashi (Graduate School of Pharmaceutical Sciences, University of Tokyo) |
|||||||||||||||||||||||
|
|||||||||||||||||||||||
| Jun. 4, 2007 | |||||||||||||||||||||||
|
|
|||||||||||||||||||||||
|
|||||||||||||||||||||||

