|
| References |
(1) |
Crocker, P. R. & Varki, A. (2001) Siglecs in the immune system. Immunology.,103, 137-45. |
|
(2) |
Sjoberg, E. R., Powell, L. D., Klein, A. & Varki, A. (1994) Natural ligands of the B cell adhesion molecule CD22 beta can be masked by 9-O-acetylation of sialic acids. J Cell Biol.,126, 549-62.
|
|
(3) |
Han, S., Collins, B. E., Bengtson, P. & Paulson, J. C. (2005) Homomultimeric complexes of CD22 in B cells revealed by protein-glycan cross-linking. Nat Chem Biol.,1, 93-97.
|
|
(4) |
Naito, Y., Takematsu, H., Koyama, S., Miyake, S., Yamamoto, H., Fujinawa, R., Sugai, M., Okuno, Y., Tsujimoto, G., Yamaji, T., Hashimoto, Y., Itohara, S., Kawasaki, T., Suzuki, A. & Kozutsumi, Y. (2007) Germinal Center Marker GL7 Probes Activation-Dependent Repression of N-Glycolylneuraminic Acid, a Sialic Acid Species Involved in the Negative Modulation of B-Cell Activation. Mol Cell Biol.,27,3008-22.
|
|
(5) |
Razi, N. & Varki, A. (1998) Masking and unmasking of the sialic acid-binding lectin activity of CD22 (Siglec-2) on B lymphocytes. Proc Natl Acad Sci USA.,95, 7469-74.
|
|
(6) |
Hennet, T., Chui, D., Paulson, J. C. & Marth, J. D. (1998) Immune regulation by the ST6Gal sialyltransferase.Proc Natl Acad Sci U S A.,95, 4504-9.
|
|
(7) |
Collins, B. E., Smith, B. A., Bengtson, P. & Paulson, J. C. (2005) Ablation of CD22 in ligand-deficient mice restores B cell receptor signaling.Nat Immunol.,7, 199-206.
|
|
(8) |
Poe, J. C., Fujimoto, Y., Hasegawa, M., Haas, K. M., Miller, A. S., Sanford, I. G., Bock, C. B., Fujimoto, M. & Tedder, T. F. (2004) CD22 regulates B lymphocyte function in vivo through both ligand-dependent and ligand-independent mechanisms. Nat Immunol.,5, 1078-87.
|
|
|
|
|
|



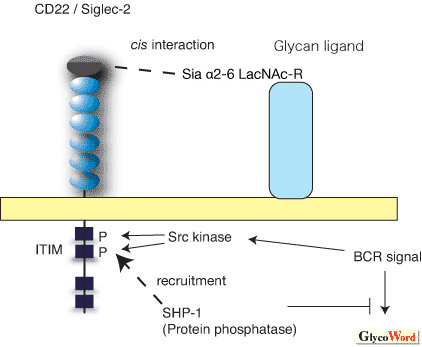
 2,6-linked sialylated glycan as a cis-ligand, which is expressed on the surface of the same cell. The intracellular cytoplasmic domain of CD22 has ITIM (immunoreceptor tyrosine inhibitory motif), which often negatively regulate the signal transduction by recruiting SH2 domain containing the tyrosine phosphatase SHP-1 upon phosphorylation. As a result, CD22 is thought to negatively regulate B cell receptor signaling (Figure 1).
2,6-linked sialylated glycan as a cis-ligand, which is expressed on the surface of the same cell. The intracellular cytoplasmic domain of CD22 has ITIM (immunoreceptor tyrosine inhibitory motif), which often negatively regulate the signal transduction by recruiting SH2 domain containing the tyrosine phosphatase SHP-1 upon phosphorylation. As a result, CD22 is thought to negatively regulate B cell receptor signaling (Figure 1).