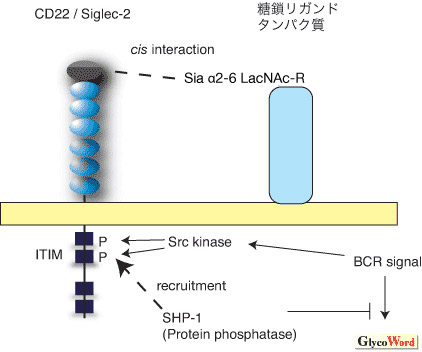
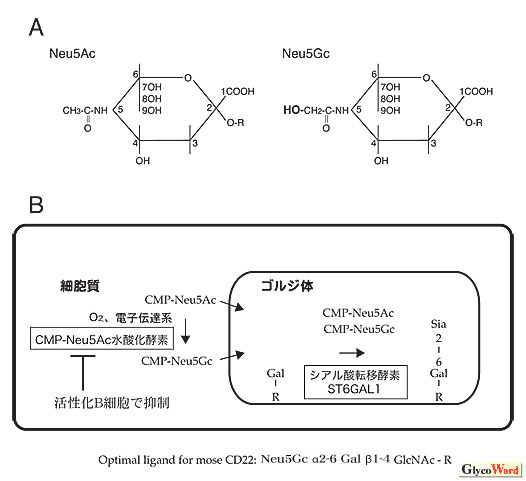
|
| References |
(1) |
Crocker, P. R. & Varki, A. (2001) Siglecs in the immune system. Immunology.,103, 137-45. |
|
(2) |
Sjoberg, E. R., Powell, L. D., Klein, A. & Varki, A. (1994) Natural ligands of the B cell adhesion molecule CD22 beta can be masked by 9-O-acetylation of sialic acids. J Cell Biol.,126, 549-62.
|
|
(3) |
Han, S., Collins, B. E., Bengtson, P. & Paulson, J. C. (2005) Homomultimeric complexes of CD22 in B cells revealed by protein-glycan cross-linking. Nat Chem Biol.,1, 93-97.
|
|
(4) |
Naito, Y., Takematsu, H., Koyama, S., Miyake, S., Yamamoto, H., Fujinawa, R., Sugai, M., Okuno, Y., Tsujimoto, G., Yamaji, T., Hashimoto, Y., Itohara, S., Kawasaki, T., Suzuki, A. & Kozutsumi, Y. (2007) Germinal Center Marker GL7 Probes Activation-Dependent Repression of N-Glycolylneuraminic Acid, a Sialic Acid Species Involved in the Negative Modulation of B-Cell Activation. Mol Cell Biol.,27,3008-22.
|
|
(5) |
Razi, N. & Varki, A. (1998) Masking and unmasking of the sialic acid-binding lectin activity of CD22 (Siglec-2) on B lymphocytes. Proc Natl Acad Sci USA.,95, 7469-74.
|
|
(6) |
Hennet, T., Chui, D., Paulson, J. C. & Marth, J. D. (1998) Immune regulation by the ST6Gal sialyltransferase.Proc Natl Acad Sci U S A.,95, 4504-9.
|
|
(7) |
Collins, B. E., Smith, B. A., Bengtson, P. & Paulson, J. C. (2005) Ablation of CD22 in ligand-deficient mice restores B cell receptor signaling.Nat Immunol.,7, 199-206.
|
|
(8) |
Poe, J. C., Fujimoto, Y., Hasegawa, M., Haas, K. M., Miller, A. S., Sanford, I. G., Bock, C. B., Fujimoto, M. & Tedder, T. F. (2004) CD22 regulates B lymphocyte function in vivo through both ligand-dependent and ligand-independent mechanisms. Nat Immunol.,5, 1078-87.
|
|
|
|
|
|