Introduction to Galectins-Experimental Protocols and Unsolved Questions (Narratives)
Oct 01, 2020
Introduction to Galectins Experimental Protocols and Unsolved Questions (Narratives) (Glycoforum. 2020 Vol.23 (5), A14) DOI: https://doi.org/10.32285/glycoforum.23A14
Jun Hirabayashi / Sachiko Sato
Jun Hirabayashi
National Institute of Advanced Industrial Science and Technology (AIST), Tsukuba, Japan. Ph.D, Science
After graduated from Tohoku University (Master of Science), he started his professional carrier at Teikyo University under Prof. Kenichi Kasai for the investigation of animal lectins. On the occasion of GlycoXV (Tokyo, 1999), he proposed the concept glycome; for this realization, he moved to National Institute of Advanced Industrial Science and Technology (AIST, Tsukuba) in 2002, and was involved in a series of national projects for glycan engineering, while he was a deputy director in Research Center for Medical Glycoscience (2006~), prime senior researcher of Research Center for Stem Cell Engineering (2012~). Now, he is a distinguished senior researcher in AIST, while being a vice president of the Japanese Society of Carbohydrate Research (JSCR) and Japanese Consortium for Glycoscience and Glycotechnology. (JCGG). He is also a visiting professor of Kagawa University (2003~) and Yokohama City University (2019~).
Sachiko Sato
Research Centre for Infectious Diseases, Faculty of Medicine, Laval University, Quebec City, Canada. Ph.D, Pharmaceutical science
Sachiko Sato graduated from Faculty of Pharmaceutical Science, Chiba University. She joined as postgraduate student in the laboratory of Dr. Akira Kobata, the Institute of Medical Science, the University of Tokyo, Japan in 1987. She also worked in the laboratory of Dr. R. Colin Hughes, MRC: National Institute for Medical Research in London, UK, where she first encountered a cytosolic mammalian lectin, now called galectin-3. She obtained her Ph. D. from the University of Tokyo in 1994. As postdoctoral fellow in the laboratory of Dr. Ron Kopito, Stanford University, she was involved in the work on cystic fibrosis. She became principal investigator of the laboratory of glycobiology in Research Center for Infectious Diseases, and assistant professor of the Faculty of Medicine, Laval University, Quebec, Canada in 1999 and is full professor since 2010. She is also director of the Bioimaging platform since 2003.
A brand new Glycoforum® genre series related to Galectins will be started, combined with publication of many detailed experimental protocols for galectin production. As editors of this series we hope to bring unique perspectives to the field of galectins and glycoscience.
Galectins are defined as an evolutionarily related family of proteins (Pfam ID: PF00337) with conserved amino acid sequences and sugar-binding specificity confined to β-galactoside 1. They are a large group of carbohydrate-binding proteins in animals and many other multicellular organisms. Galectins have been investigated over a long period (Fig. 1) and found to participate in an extensive range of biological processes, such as carcinogenesis, immunity, development, infections, etc. (see Atlas of galectin in Fig. 2; modified from the original figure from ref. 2). Despite the long history of galectin research, many unsolved mysteries remain: Prof. Kasai once referred to these mysteries as the “Seven wonders of galectins”3: namely, 1) What is the general mission of galectins? 2) Why are they extracellularly located in spite of the fact that they were designed to be intracellular proteins? 3) How do cells externalize them? 4) Why are all galectins galactoside-specific? 5) What are their extracellular and intracellular ligands? 6) Why are they localized at sites where the risk of oxidative inactivation is high? 7) Why do galectins and legume lectins have a common topology? It is simply surprising that these questions were raised almost a quarter century ago, but none of them have been answered until today.
Fig. 1. Epoch-making events in the history of galectin research. Based on the judgment of the editors (J.H. and S.S.), the focus was selected to be relatively early investigations.
* This classification of galectin names is based on the observation that the galectins studied in the early days of galectin research lost their carbohydrate-binding activity in the absence of thiol (SH)-reducing reagents, such as 2-mercaptoethanol, but this naming turned out later to be not valid for many other members (nowadays, it is understood that readily oxidatively-inactivated galectin-1 is exceptional). This confusion was one of the reasons for redefinition of the systematic name “galectin” by a group of researchers in 1994, led by SH Barondes.
References (in part of Fig. 1)
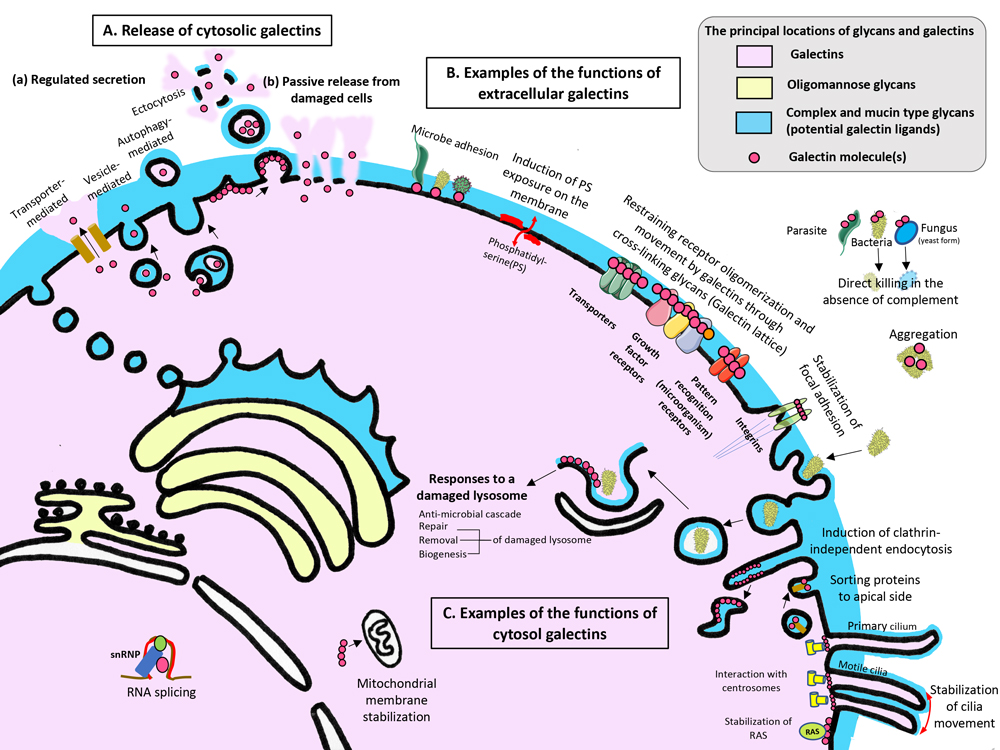
Figure 2. An atlas of galectins.
The principal locations of galectins (light pink), their ligands, complex and mucin type glycans (light blue) and oligomannose type glycans (light yellow) are shown. Galectin molecules are depicted as pink dots.
A: Cytosolic galectins can be released into the extracellular milieu through regulated secretion (a) and passive release from damaged cells (b). In the case of regulated secretion (a), galectins are known to be secreted using four pathways: transporter-, vesicle-, and autophagy-mediated and ectocytosis pathways.
B: Once released to the extracellular milieu, galectins exert many functions in a spatiotemporal and context-dependent manner. Most of those functions rely on their glycan binding activity and multivalency in their binding to glycans. Some of those functions are illustrated here.
C: Within the cytosol, galectins mediate various responses through glycan-galectin and/or protein-galectin interactions. Some of those functions are illustrated here.
For a detailed explanation, please refer to either the Glycoforum article by S. Sato or the reference (S. Sato, Mammalian cytosolic galectins act as damage-associated molecular patterns, resolutions-associated molecular patterns, and pattern-recognition receptors in innate immunity, Comprehensive Glycoscience 2nd Edition in press).
In the same paper, however, he gave a unique, witty answer to the first question, i.e., “Galectins are intelligent glue as well as non-bureaucratic bureaucrats or almighty supporting actors”. Questions 2, 3, 5, and 6 are about the binary roles of galectins in both cytoplasmic and extracellular spaces. Probably the most essential question, if combining them, is on the “origin” of galectins. To make a long history short, this is the ultimate, chicken or egg-type question; that is, “Were galectin ancestors born as lectins or non-lectin proteins?” Considering that none of the galectins identified so far have a signal sequence for secretion and none of them are glycosylated, their ancestors must have originated as cytoplasmic proteins essentially without glycans, but their function(s) as lectins in cytoplasm remained elusive until recently.
The remaining two, i.e., questions 4 and 7, are about the protein scaffold (β-sandwich/jellyroll) and carbohydrate-specificity of galectins. In other words, another essential question is “Why is galectins’ specificity confined to β-galactoside?”, but nobody seems to have found an a priori answer to it. This is again an issue of galectins evolution, but can we find any useful structural clue to answer this question?
The Galectins series consists of two parts: Part I is Experimental protocols for galectin expression. Experience-based detailed established protocols are kindly provided by Nozomu Nishi (Kagawa University, Japan). As many galectin researchers know, he has long been involved in basic researches including studies on the production of a series of galectins (mainly of human origin) and their useful mutants, including the stabilized cysteine-less Gal-1 mutant (designated CSGal-1) 4 and linker-deleted Gal-8 and Gal-9 mutants (G8Null, G9Null) 5. He also intended to deliver a wide range of expression plasmids through a public bio-resource center at RIKEN (Wako, Japan). His detailed protocol is not only for those who have been working on recombinant lectins but also for researchers including young scientists, who are not familiar with lectins purification and find the biochemical protocol currently used to produce recombinant proteins, extremely challenging, even with the fully established Escherichia coli system. It is true that there are many pitfalls, the details of which are not necessarily described in the original research papers. The provided experimental protocols consist of 1) a “27-step experimental protocol” with helpful notes, 2) “comments referring to individual human galectins”, and 3) “supplementary material” including other related experimental methods (e.g., how to concentrate galectin samples before SDS-PAGE) and specific comments (e.g., on how to purify Gal-9 under acidic conditions).
Part II is a series of Narratives, entitled “Unsolved Questions about Galectins”, which focus on some of the important unsolved questions mentioned above, from an evolutionary perspective. For this series, we invited a few world-renowned scientists who have worked on galectins or galectin-related issues for many years. They provide attractive topics, sometimes giving us bold hypotheses and exciting imaginative speculation, which would be difficult to publish in conventional scientific literature. Indeed, it is our very intention to initiate this series and by doing so, create opportunities to discuss their views and perspectives on galectins from different angles. As far as we know, no such attempt has been made, while several successful special issues on galectins have been published6-9. We greatly thank the following authors for their understanding of our planning concept and kindly accepting our invitation to write about their own interests (the order of publication may change, and additional author(s) might join in this forum). As editors of this series, we hope to bring unique perspectives into the field of galectins and glycoscience.
■ Kenichi Kasai (Teikyo University, Japan)
■ Hans-Joachim Gabius (Ludwig-Maximilians-University Munich, Germany)
■ Hakon Leffler (Lund University, Sweden)
■ James W. Dennis (Mount Sinai Hospital, Toronto, Canada)
■ Tadasu Urashima (Obihiro University of Agriculture and Veterinary Medicine, Obihiro, Japan)
■ Sachiko Sato (Laval University, Quebec City, Canada)
■ Jun Hirabayashi (AIST, Tsukuba, Japan)
References
Barondes SH, Castronovo V, Cooper DN, Cummings RD, Drickamer K, Feizi T, Gitt MA, Hirabayashi J, Hughes C, Kasai K, Leffler H, Liu F-T, Lotan R, Mercurio AM, Monsigny M, Pillai S, Poirer F, Raz A, Rigby PWJ, Wang JL (1994) Galectins: a family of animal beta-galactoside-binding lectins. Cell. 76, 597–8.
Sato S (2020) Galectins and innate immunity, Comprehensive Glycoscience 2nd edition Vol 5 Chapter 147. In press
Kasai K (1997) Galectin: intelligent glue, non-bureaucratic bureaucrat or almighty supporting actor. Trends Glycosic Glycotechnol. 9(45), 167-70.
Nishi N, Abe A, Iwaki J, Yoshida H, Itoh A, Shoji H, Kamitori S, Hirabayashi J, Nakamura T (2008) Functional and structural bases of a cysteine-less mutant as a long-lasting substitute for galectin-1. Glycobiology. 18(12), 1065-73.
Nishi N, Itoh A, Fujiyama A, Yoshida N, Araya S, Hirashima M, Shoji H, Nakamura T (2005) Development of highly stable galectins: truncation of the linker peptide confers protease-resistance on tandem-repeat type galectins. FEBS Lett. 579(10), 2058-64.
Kilpatrick DC (2002) Animal lectins: a historical introduction and overview. Biochim Biophys Acta. 1572, 187–97.
Barondes SH (1997) Galectins: A personal overview. Trends Glycosci Glycotechnol. 9, 1–7.
Reitherman R W, Rosen SD, Frasier WA, Barondes SH (1975) Cell surface species-specific high affinity receptors for discoidin: developmental regulation in Dictyostelium discoideum. Proc Natl Acad Sci U S A. 72, 3541–5.
Teichberg VI, Silman I, Beitsch DD, Resheff G (1975) A beta-D-galactoside binding protein from electric organ tissue of Electrophorus electricus. Proc Natl Acad Sci U S A. 72, 1383–7.
Leffler H, Barondes SH (1986) Specificity of binding of three soluble rat lung lectins to substituted and unsubstituted mammalian b-galactosides. J Biol Chem. 261, 10119–26.
Ohyama Y, Hirabayashi J, Oda Y, Ohno S, Kawasaki H, Suzuki K, Kasai K (1986) Nucleotide sequence of chick 14K beta-galactoside-binding lectin mRNA. Biochem Biophys Res Commun. 134, 51–6.
Drickamer K (1988) Two distinct classes of carbohydrate-recognition domains in animal lectins. J Biol Chem. 263, 9557–60.
Cooper DN, Barondes SH (1990) Evidence for export of a muscle lectin from cytosol to extracellular matrix and for a novel secretory mechanism. J Cell Biol. 110, 1681–91.
Hirabayashi J, Satoh M, Kasai K (1992) Evidence that Caenorhabditis elegans 32-kDa beta-galactoside-binding protein is homologous to vertebrate beta-galactoside-binding lectins. cDNA cloning and deduced amino acid sequence. J Biol Chem. 267, 15485–90.
Hirabayashi J, Kasai K (1993) The family of metazoan metal-independent beta-galactoside-binding lectins: structure, function and molecular evolution. Glycobiology. 3, 297–304.
Lobsanov YD, Gitt MA, Leffler H, Barondes SH, Rini JM (1993) X-ray crystal structure of the human dimeric S-Lac lectin, L-14-II, in complex with lactose at 2.9-A resolution. J Biol Chem. 268, 27034–8.
Poirier F, Robertson EJ (1993) Normal development of mice carrying a null mutation in the gene encoding the L14 S-type lectin. Development. 119, 1229–36.
Barondes SH, Castronovo V, Cooper DN, Cummings RD, Drickamer K, Feizi T, Gitt MA, Hirabayashi J, Hughes C, Kasai K et al (1994) Galectins: a family of animal beta-galactoside-binding lectins. Cell. 76, 597–8.
Dagher SF, Wang JL, Patterson RJ (1995) Identification of galectin-3 as a factor in pre-mRAN splicing. Proc Natl Acad Sci U S A. 92, 1213–1217
Cooper DN, Boulianne RP, Charlton S, Farrell EM, Sucher A, Lu BC (1997) Fungal galectins, sequence and specificity of two isolectins from Coprinus cinereus. J Biol Chem. 272, 1514–21.
Matsumoto R, Matsumoto H, Seki M, Hata M, Asano Y, Kanegasaki S, Stevens RL, Hirashima M (1998) Human ecalectin, a variant of human galectin-9, is a novel eosinophil chemoattractant produced by T lymphocytes. J Biol Chem. 273, 16976–84.
Cooper DN, Barondes SH (1999) God must love galectins; he made so many of them. Glycobiology. 9, 979–84.
Hughes RC (1999) Secretion of the galectin family of mammalian carbohydrate-binding proteins. Biochim Biophys Acta. 1473, 172–85.
Demetriou M, Granovsky M, Quaggin S, Dennis JW (2001) Negative regulation of T-cell activation and autoimmunity by Mgat5 N-glycosylation. Nature. 409, 733–9.
Hirabayashi J, Hashidate, T, Arata Y, Nishi N, Nakamura T, Hirashima M, Urashima T, Oka T, Futai M, Muller WE, YagiF, Kasai K. (2002) Oligosaccharide specificity of galectins: a search by frontal affinity chromatography. Biochim Biophys Acta. 1572, 232–54.
Sasai K, Ikeda Y, Fujii T, Tsuda T, Taniguchi N (2002) UDP-GlcNAc concentration is an important factor in the biosynthesis of beta1,6-branched oligosaccharides: regulation based on the kinetic properties of N-acetylglucosaminyltransferase V. Glycobiology. 12, 119–27.
Sato S (2002) Galectin as a molecule of danger signal, which could evoke immune response to infection. Trends Glycosci Glycotechnol. 14, 285–301
Sato S, Nieminen J (2004) Seeing strangers or announcing “danger”: Galectin-3 in two models of innate immunity. Glycoconj J. 19, 583–91.
Delacour D, Gouyer V, Zanetta JP, Drobecq H, Leteurtre E, Grard G, Moreau-Hannedouche O, Maes E, Pons A, Andre S, Le Bivic A, Gabius HJ, Manninen A, Simons K, Huet G (2005) Galectin-4 and sulfatides in apical membrane trafficking in enterocyte-like cells. J Cell Biol. 169, 491–501
Kohatsu L, Hsu DK, Jegalian AG, Liu FT, Baum LG (2006) Galectin-3 induces death of Candida species expressing specific beta-1,2-linked mannans. J Immunol. 177, 4718–26.
Stowell SR, Karmakar S, Stowell CJ, Dias-Baruffi M, McEver RP, Cummings RD (2007) Human galectin-1, -2, and -4 induce surface exposure of phosphatidylserine in activated human neutrophils but not in activated T cells. Blood. 109, 219–27.
Nieminen J, Kuno A, Hirabayashi J, Sato S (2007) Visualization of galectin-3 oligomerization on the surface of neutrophils and endothelial cells using fluorescence resonance energy transfer. J Biol Chem. 282, 1374–83.
Lau KS, Partridge EA, Grigorian A, Silvescu CI, Reinhold VN, Demetriou M, Dennis JW (2007) Complex N-glycan number and degree of branching cooperate to regulate cell proliferation and differentiation. Cell. 129, 123–34.
Paz I, Sachse M, Dupont N, Mounier J, Cederfur C, Enninga J, Leffler H, Poirier F, Prevost MC, Lafont F, Sansonetti P (2009) Galectin-3, a marker for vacuole lysis by invasive pathogens. Cell Microbiol. 12, 530–44.
Dupont N, Lacas-Gervais S, Bertout J, Paz I, Freche B, Van Nhieu GT, van der Goot FG, Sansonetti PJ, Lafont F (2009) Shigella phagocytic vacuolar membrane remnants participate in the cellular response to pathogen invasion and are regulated by autophagy. Cell Host Microbe. 6, 137–49.
Thurston TL, Wandel MP, Muhlinen von N, Foeglein A, Randow F (2012) Galectin 8 targets damaged vesicles for autophagy to defend cells against bacterial invasion. Nature. 482, 414–8.
MacKinnon AC, Gibbons MA, Farnworth SL, Leffler H, Nilsson UJ, Delaine T, Simpson AJ, Forbes SJ, Hirani N, Gauldie J, Sethi T (2012) Regulation of transforming growth factor-β1-driven lung fibrosis by galectin-3. Am J Resp Crit Care Med. 185, 537–46.
Lin Y-H, Qiu D-C, Chang W-H, Yeh Y-Q, Jeng U-S, Liu F-T, Huang J-R (2017) The intrinsically disordered N-terminal domain of galectin-3 dynamically mediates multisite self-association of the protein through fuzzy interactions. J Biol Chem. 292, 17845–56.
Ludwig A-K, Michalak M, Xiao Q, Gilles U, Medrano FJ, Ma H, FitzGerald FG, Hasley WD, Melendez-Davila A, Liu M, Rahimi K, Kostina NY, Rodriguez-Emmenegger C, Möller M, Lindner I, Kaltner H, Cudic M, Reusch D, Kopitz J, Romero A, Oscarson S, Klein ML, Gabius H-J, Percec V (2019) Design-functionality relationships for adhesion/growth-regulatory galectins. Proc Natl Acad Sci U S A. 116, 2837–42.