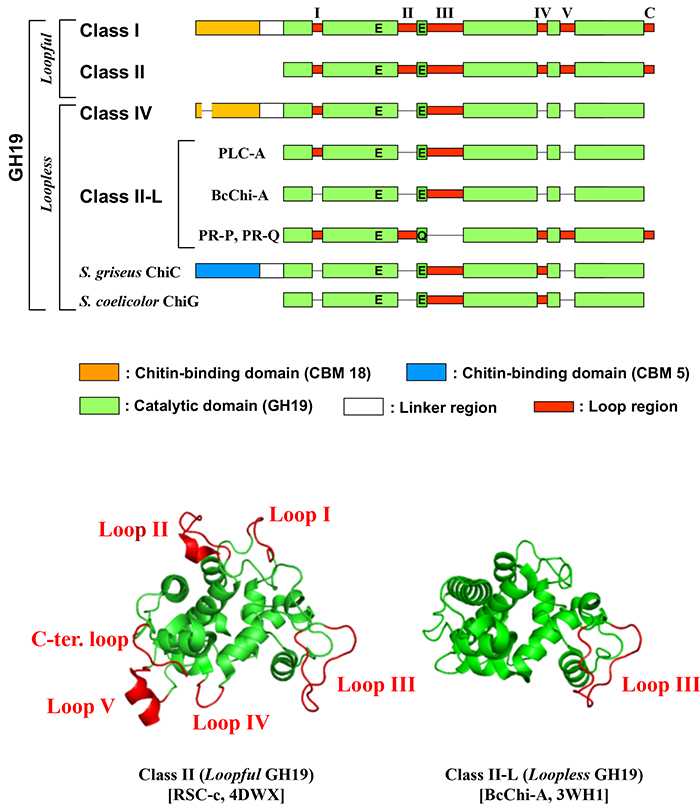
GH19の触媒ドメインは、最初に立体構造が明らかとなったオオムギ由来GH19キチナーゼの構造に基づき、N末端より5つの内部ループ領域とC末端ループ領域の合計6つのループ領域が存在することが分かっている(図 1)6。GH19キチナーゼにおいて、このループ領域が全てあるものと、一部欠損しているものがあり、欠損の数や場所にはバリエーションがある.アメリカヤマゴボウ緑葉由来キチナーゼ-A(PLC-A)7はloop II, IV, VおよびC末端loopが、ナガハハリガネゴケ由来キチナーゼ-A(BcChi-A)8,9はloop III以外のループ領域が、タバコ由来の感染特異的発現タンパク質(Pathogenesis-related Protein: PR-protein)であるPR-PとPR-Q10はloop IIIのみが欠失している(図 1)。細菌類由来GH19キチナーゼであるStreptomyces griseus由来キチナーゼ-C11およびStreptomyces coelicolor由来キチナーゼ-G12は何れもloop I, II, VおよびC末端loopが欠失している(図 2)。
図 1. GH19キチナーゼのクラス分類と構造特性 上段はGH19キチナーゼの4つのクラス(I, II, IV, II-L)の模式図.ループ領域の上に付したI-VとC はループ名を表す。 PLC-A, pokeweed leaf chitinase-A (Q7M1Q9); BcChi-A, Bryum coronatum chitinase-A (BAF99002); PR-P and PR-Q, pathogenesis-related protein P and Q from Nicotiana tabacum (CAA35790 and CAA35789), S. griseus 由来GH19 Chitinase-C (BAA23739); Chi-G, S. coelicolor 由来GH19キチナーゼ-G (CAD55444). 下段はGH19キチナーゼのリボンモデル。各モデルの下の2行目中括弧内右はPDB IDを示す。
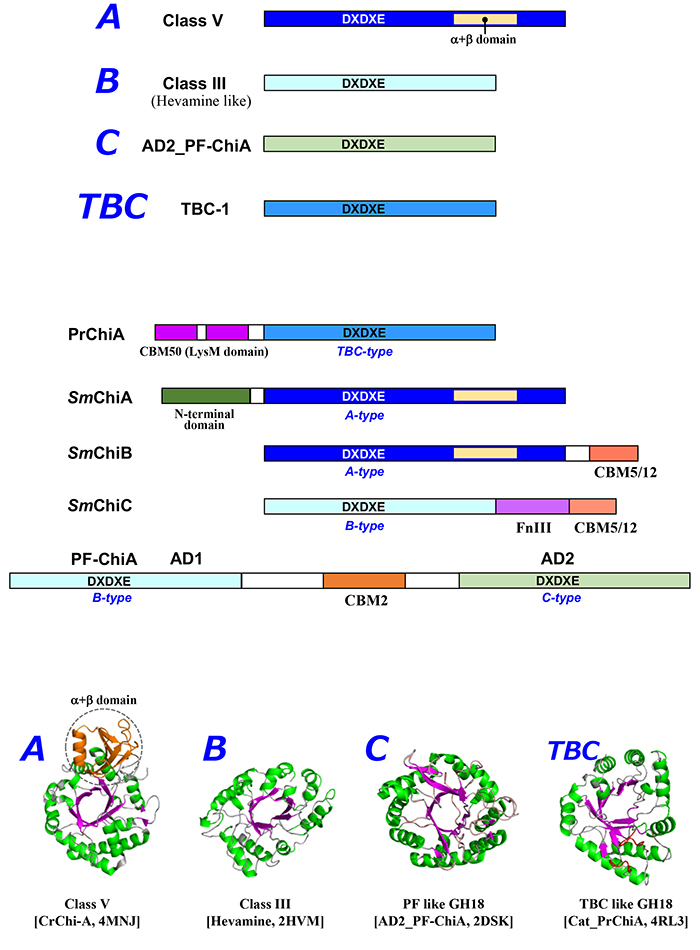
図 2. GH18キチナーゼの分類と構造特性 上段はGH18のAタイプ、Bタイプ、CタイプおよびTBCタイプの模式図。中段はGH18触媒ドメインを含む代表的なキチナーゼのドメイン構成の模式図。PrChiA, Pteris ryukyuensis chitinase-A (BAE98134.1); SmChiA: Serratia marcescens Chitinase A (BAA31567.1); SmChiA, S. marcescens Chitinase B (CAA85292.1); SmChiA, S. marcescens Chitinase C (CAF74787.1); PF-ChiA, Pyrococcus furiosus putative chitinase (AAL81357.1) 。下段はGH18キチナーゼのリボンモデルを示す。
Theis T, Stahl U. Antifungal proteins: targets, mechanisms and prospective applications. Cell Mol Life Sci. 2004 Feb;61(4):437-55. doi: 10.1007/s00018-003-3231-4.
Watanabe T, Kanai R, Kawase T, Tanabe T, Mitsutomi M, Sakuda S, Miyashita K. Family 19 chitinases of Streptomyces species: characterization and distribution. Microbiology (Reading). 1999 Dec;145 (Pt 12):3353-3363. doi: 10.1099/00221287-145-12-3353.
Kawase T, Yokokawa S, Saito A, Fujii T, Nikaidou N, Miyashita K, Watanabe T. Comparison of enzymatic and antifungal properties between family 18 and 19 chitinases from S. coelicolor A3(2).
Drula E, Garron ML, Dogan S, Lombard V, Henrissat B, Terrapon N. The carbohydrate-active enzyme database: functions and literature. Nucleic Acids Res. 2022 Jan 7;50(D1):D571-D7. doi: 10.1093/nar/gkab1045.
Hart PJ, Monzingo AF, Ready MP, Ernst SR, Robertus JD. Crystal structure of an endochitinase from Hordeum vulgare L. seeds. J Mol Biol. 1993 Jan 5;229(1):189-93. doi: 10.1006/jmbi.1993.1017.
Yamagami T, Tanigawa M, Ishiguro M, Funatsu G. Complete amino acid sequence of chitinase-A from leaves of pokeweed (Phytolacca americana). Biosci Biotechnol Biochem. 1998 Apr;62(4):825-8. doi: 10.1271/bbb.62.825.
Taira T, Mahoe Y, Kawamoto N, Onaga S, Iwasaki H, Ohnuma T, Fukamizo T. Cloning and characterization of a small family 19 chitinase from moss (Bryum coronatum). Glycobiology. 2011 May;21(5):644-54. doi: 10.1093/glycob/cwq212.
Ohnuma T, Umemoto N, Nagata T, Shinya S, Numata T, Taira T, Fukamizo T. Crystal structure of a "loopless" GH19 chitinase in complex with chitin tetrasaccharide spanning the catalytic center. Biochim Biophys Acta. 2014 Apr; 1844(4):793-802. doi: 10.1016/j.bbapap.2014.02.013.
Payne G, Ahl P, Moyer M, Harper A, Beck J, Meins F Jr, Ryals J. Isolation of complementary DNA clones encoding pathogenesis-related proteins P and Q, two acidic chitinases from tobacco. Proc Natl Acad Sci U S A. 1990 Jan; 87(1):98-102. doi: 10.1073/pnas.87.1.98.
Kezuka Y, Ohishi M, Itoh Y, Watanabe J, Mitsutomi M, Watanabe T, Nonaka T. Structural studies of a two-domain chitinase from Streptomyces griseus HUT6037. J Mol Biol. 2006 Apr 28; 358(2):472-84. doi: 10.1016/j.jmb.2006.02.013.
Hoell IA, Dalhus B, Heggset EB, Aspmo SI, Eijsink VG. Crystal structure and enzymatic properties of a bacterial family 19 chitinase reveal differences from plant enzymes. FEBS J. 2006 Nov; 273(21):4889-900. doi: 10.1111/j.1742-4658.2006.05487.x.
Ohnuma T, Umemoto N, Kondo K, Numata T, Fukamizo T. Complete subsite mapping of a "loopful" GH19 chitinase from rye seeds based on its crystal structure. FEBS Lett. 2013 Aug 19; 587(16):2691-7. doi: 10.1016/j.febslet.2013.07.008.
Broekaert I, Lee HI, Kush A, Chua NH, Raikhel N. Wound-induced accumulation of mRNA containing a hevein sequence in laticifers of rubber tree (Hevea brasiliensis). Proc Natl Acad Sci U S A. 1990 Oct; 87(19):7633-7. doi: 10.1073/pnas.87.19.7633.
Shinshi H, Neuhas JM, Ryals J, Meins F Jr. Structure of a tobacco endochitinase gene: evidence that different chitinase genes can arise by transposition of sequences encoding a cysteine-rich domain. Plant Mol Biol. 1990 Mar; 14(3):357-68. doi: 10.1007/BF00028772.
Yano S, Suyotha W, Honda A, Takagi K, Rattanakit-Chandet N, Wakayama M, Tachiki T. N-terminal region of chitinase I of Bacillus circulans KA-304 contained new chitin-biding domain. Biosci Biotechnol Biochem. 2011; 75(2):299-304. doi: 10.1271/bbb.100659.
Collinge DB, Kragh KM, Mikkelsen JD, Nielsen KK, Rasmussen U, Vad K. Plant chitinases. Plant J. 1993 Jan; 3(1):31-40. doi: 10.1046/j.1365-313x.1993.t01-1-00999.x.
Terwisscha van Scheltinga AC, Hennig M, Dijkstra BW. The 1.8 A resolution structure of hevamine, a plant chitinase/lysozyme, and analysis of the conserved sequence and structure motifs of glycosyl hydrolase family 18. J Mol Biol. 1996 Sep 20; 262(2):243-57. doi: 10.1006/jmbi.1996.0510.
Yamagami T, Ishiguro M. Complete amino acid sequences of chitinase-1 and -2 from bulbs of genus Tulipa. Biosci Biotechnol Biochem. 1998 Jun; 62(6):1253-7. doi: 10.1271/bbb.62.1253.
Onaga S, Taira T. A new type of plant chitinase containing LysM domains from a fern (Pteris ryukyuensis): roles of LysM domains in chitin binding and antifungal activity. Glycobiology. 2008 May;18(5):414-23. doi: 10.1093/glycob/cwn018.
Kitaoku Y, Umemoto N, Ohnuma T, Numata T, Taira T, Sakuda S, Fukamizo T. A class III chitinase without disulfide bonds from the fern, Pteris ryukyuensis: crystal structure and ligand-binding studies. Planta. 2015 Oct;242(4):895-907. doi: 10.1007/s00425-015-2330-4.
Ohnuma T, Numata T, Osawa T, Mizuhara M, Vårum KM, Fukamizo T. Crystal structure and mode of action of a class V chitinase from Nicotiana tabacum. Plant Mol Biol. 2011 Feb;75(3):291-304. doi: 10.1007/s11103-010-9727-z.
Melchers LS, Apotheker-de Groot M, van der Knaap JA, Ponstein AS, Sela-Buurlage MB, Bol JF, Cornelissen BJ, van den Elzen PJ, Linthorst HJ. A new class of tobacco chitinases homologous to bacterial exo-chitinases displays antifungal activity. Plant J. 1994 Apr;5(4):469-80. doi: 10.1046/j.1365-313x.1994.5040469.x.
Suzuki K, Taiyoji M, Sugawara N, Nikaidou N, Henrissat B, Watanabe T. The third chitinase gene (chiC) of Serratia marcescens 2170 and the relationship of its product to other bacterial chitinases. Biochem J. 1999 Nov 1;343 Pt 3(Pt 3):587-96.
Seidl V, Huemer B, Seiboth B, Kubicek CP. A complete survey of Trichoderma chitinases reveals three distinct subgroups of family 18 chitinases. FEBS J. 2005 Nov;272(22):5923-39. doi: 10.1111/j.1742-4658.2005.04994.x.
Nakamura T, Mine S, Hagihara Y, Ishikawa K, Uegaki K. Structure of the catalytic domain of the hyperthermophilic chitinase from Pyrococcus furiosus. Acta Crystallogr Sect F Struct Biol Cryst Commun. 2007 Jan 1;63(Pt 1):7-11. doi: 10.1107/S1744309106051773.
Taira T, Gushiken C, Sugata K, Ohnuma T, Fukamizo T. Unique GH18 chitinase from Euglena gracilis: full-length cDNA cloning and characterization of its catalytic domain. Biosci Biotechnol Biochem. 2018 Jul;82(7):1090-1100. doi: 10.1080/09168451.2018.1459463.
Wang J, Chitsaz F, Derbyshire MK, Gonzales NR, Gwadz M, Lu S, Marchler GH, Song JS, Thanki N, Yamashita RA, Yang M, Zhang D, Zheng C, Lanczycki CJ, Marchler-Bauer A. The conserved domain database in 2023. Nucleic Acids Res. 2023 Jan 6;51(D1):D384-D388. doi: 10.1093/nar/gkac1096.
Schlumbaum, A., Mauch, F., Vögeli, U. et al. Plant chitinases are potent inhibitors of fungal growth. Nature. 324, 365–367 (1986).
Toki Taira. Structures and antifungal activity of plant chitinases. J Appl Glycosci. 2010 Aug;57(3):167-176. doi: 10.5458/jag.57.167
Ohnuma T, Taira T, Fukamizo T: Antifungal activity of recombinant class V chitinases from Nicotiana tabacum and Arabidopsis thaliana. J Appl Glycosci. 2012 Jan 59(1):47-50. doi: 10.5458/jag.jag.JAG-2011_019
Garcia-Casado G, Collada C, Allona I, Casado R, Pacios LF, Aragoncillo C, Gomez L. Site-directed mutagenesis of active site residues in a class I endochitinase from chestnut seeds. Glycobiology. 1998 Oct;8(10):1021-8. doi: 10.1093/glycob/8.10.1021.
Andersen MD, Jensen A, Robertus JD, Leah R, Skriver K. Heterologous expression and characterization of wild-type and mutant forms of a 26 kDa endochitinase from barley (Hordeum vulgare L.). Biochem J. 1997 Mar 15;322 ( Pt 3)(Pt 3):815-22. doi: 10.1042/bj3220815.
Ohnuma T, Taira T, Yamagami T, Aso Y, Ishiguro M. Molecular cloning, functional expression, and mutagenesis of cDNA encoding class I chitinase from rye (Secale cereale) seeds. Biosci Biotechnol Biochem. 2004 Feb;68(2):324-32. doi: 10.1271/bbb.68.324.
Takashima T, Henna H, Kozome D, Kitajima S, Uechi K, Taira T. cDNA cloning, expression, and antifungal activity of chitinase from Ficus microcarpa latex: difference in antifungal action of chitinase with and without chitin-binding domain. Planta. 2021 May 13;253(6):120. doi: 10.1007/s00425-021-03639-8.
Broekaert WF, Mariën W, Terras FR, De Bolle MF, Proost P, Van Damme J, Dillen L, Claeys M, Rees SB, Vanderleyden J, Cammue BPA. Antimicrobial peptides from Amaranthus caudatus seeds with sequence homology to the cysteine/glycine-rich domain of chitin-binding proteins. Biochemistry. 1992 May 5;31(17):4308-14. doi: 10.1021/bi00132a023.
Taira T, Ohnuma T, Yamagami T, Aso Y, Ishiguro M, Ishihara M. Antifungal activity of rye (Secale cereale) seed chitinases: the different binding manner of class I and class II chitinases to the fungal cell walls. Biosci Biotechnol Biochem. 2002 May;66(5):970-7. doi: 10.1271/bbb.66.970.
Jacobsen, S., Mikkelsen, J.D. and Hejgaard, J. Characterization of two antifungal endochitinases from barley grain. Physiologia Plantarum. 1990 Jul;79:554-562. doi.org/10.1111/j.1399-3054.1990.tb02117.x
Iseli B, Boller T, Neuhaus JM. The N-terminal cysteine-rich domain of tobacco class I chitinase is essential for chitin binding but not for catalytic or antifungal activity. Plant Physiol. 1993 Sep;103(1):221-6. doi: 10.1104/pp.103.1.221.
Suarez V, Staehelin C, Arango R, Holtorf H, Hofsteenge J, Meins F Jr. Substrate specificity and antifungal activity of recombinant tobacco class I chitinases. Plant Mol Biol. 2001 Mar;45(5):609-18. doi: 10.1023/a:1010619421524.
Taira T, Yamagami T, Aso Y, Ishiguro M, Ishihara M. Localization, accumulation, and antifungal activity of chitinases in rye (Secale cereale) seed. Biosci Biotechnol Biochem. 2001 Dec;65(12):2710-8. doi: 10.1271/bbb.65.2710.
Itoh Y, Watanabe J, Fukada H, Mizuno R, Kezuka Y, Nonaka T, Watanabe T. Importance of Trp59 and Trp60 in chitin-binding, hydrolytic, and antifungal activities of Streptomyces griseus chitinase C. Appl Microbiol Biotechnol. 2006 Oct;72(6):1176-84. doi: 10.1007/s00253-006-0405-7.
Tsujibo H, Kubota T, Yamamoto M, Miyamoto K, Inamori Y. Characterization of chitinase genes from an alkaliphilic actinomycete, Nocardiopsis prasina OPC-131. Appl Environ Microbiol. 2003 Feb;69(2):894-900. doi: 10.1128/AEM.69.2.894-900.2003.
Taira T, Toma N, Ishihara M. Purification, characterization, and antifungal activity of chitinases from pineapple (Ananas comosus) leaf. Biosci Biotechnol Biochem. 2005 Jan;69(1):189-96. doi: 10.1271/bbb.69.189.
Taira T, Ohdomari A, Nakama N, Shimoji M, Ishihara M. Characterization and antifungal activity of gazyumaru (Ficus microcarpa) latex chitinases: both the chitin-binding and the antifungal activities of class I chitinase are reinforced with increasing ionic strength. Biosci Biotechnol Biochem. 2005 Apr;69(4):811-8. doi: 10.1271/bbb.69.811.
Takashima T, Henna H, Kozome D, Kitajima S, Uechi K, Taira T. cDNA cloning, expression, and antifungal activity of chitinase from Ficus microcarpa latex: difference in antifungal action of chitinase with and without chitin-binding domain. Planta. 2021 May 13;253(6):120. doi: 10.1007/s00425-021-03639-8.
Kang X, Kirui A, Muszyński A, Widanage MCD, Chen A, Azadi P, Wang P, Mentink-Vigier F, Wang T. Molecular architecture of fungal cell walls revealed by solid-state NMR. Nat Commun. 2018 Jul 16;9(1):2747. doi: 10.1038/s41467-018-05199-0.
Fujikawa T, Kuga Y, Yano S, Yoshimi A, Tachiki T, Abe K, Nishimura M. Dynamics of cell wall components of Magnaporthe grisea during infectious structure development. Mol Microbiol. 2009 Aug;73(4):553-70. doi: 10.1111/j.1365-2958.2009.06786.x.
Yano S, Wakayama M, Tachiki T. Cloning and expression of an alpha-1,3-glucanase gene from Bacillus circulans KA-304: the enzyme participates in protoplast formation of Schizophyllum commune. Biosci Biotechnol Biochem. 2006 Jul;70(7):1754-63. doi: 10.1271/bbb.60095.
Wu H, Shimoi H, Ito K. Purification and characterization of beta-1,6-glucanase of Streptomyces rochei application in the study of yeast cell wall proteins. Biosci Biotechnol Biochem. 2002 Nov;66(11):2515-9. doi: 10.1271/bbb.66.2515.
Tamura K., Stecher G., and Kumar S. MEGA 11: Molecular Evolutionary Genetics Analysis Version 11. Molecular Biology and Evolution. 2021. doi: 10.1093/molbev/msab120.
Le S.Q. and Gascuel O. An Improved General Amino Acid Replacement Matrix. Mol Biol Evol. 2008;25(7):1307-1320. doi: 10.1093/molbev/msn067