
Scott Strong
Scott Strong is a colorectal surgeon at the Cleveland Clinic Foundation in Cleveland, Ohio, and holds a joint appointment in the Lerner Research Institute, Department of Immunology. He received his B.S. degree in chemical engineering from Iowa State University, and his M.D. degree from the University of Iowa. He subsequently completed a surgical residency at Michigan State University - Butterworth Hospital prior to beginning his colorectal surgery training at the Cleveland Clinic in 1990. In addition, he did a research fellowship with Dr. Claudio Fiocchi, an established leader in the field of inflammatory bowel disease research. When he was appointed to his current position in 1993, Dr. Strong also established a research laboratory dedicated to investigating the cellular mechanisms of the pathogenesis of inflammatory bowel disease. His research emphasizes immune cell / non-immune cell interactions of the intestinal mucosa and has recently focused on a novel, virus-induced mechanism of cell interaction mediated by hyaluronan and CD44.

Carol de la Motte
Carol de la Motte received her B.S. degree from Cornell University, Ithaca, N.Y., where she majored in microbiology. She pursued graduate education in medical microbiology, specializing in medical mycology, at the University of Kentucky in Lexington. In 1978, she joined the Research Division of the Cleveland Clinic, where her long standing interest in factors that affect immune cell interaction with non-immune cells began. In the laboratory of Dr. Max Proffitt, she investigated virus-lymphocyte interaction, and how it affected immune cell killing of mesenchymal cells. Subsequently she moved to Dr. Paul DiCorleto's laboratory, where she became interested in leukocyte adhesion molecules expressed by endothelial cells and their implications in atherosclerosis and inflammatory diseases. Several years ago, she joined Dr. Scott Strong's laboratory, in the Department of Colorectal Surgery, to investigate cellular mechanisms of mesenchymal cell participation in inflammatory bowel disease. Serendipitously, the investigation led them to hyaluronan, and a wonderfully enjoyable, productive collaboration with Dr. Vincent Hascall and his laboratory.
Introduction Homeostasis is a state of physiologic equilibrium maintained by a balance of chemical composition and functions in coordinated organ systems. The specialized function of an individual organ requires its myriad cells of varying embryological origin to act in concert. The orchestrated cellular activities that enable homeostasis in higher organisms are staggering in number and complexity. A previous article describes some essential roles of hyaluronan in the morphogenic organization of a variety of organs, as well as its role in maintaining tissue function. a
a See review by Toole in this series.
The cell groups within tissues are organized in a manner that allows effective responses to alterations within the environment (e.g., physical damage, ingested or inhaled chemicals, temperature changes, and microbial attacks) that protect the organism when necessary followed by a return to basal, tissue-maintenance activity. To meet the needs of the individual organ as well as the entire organism, cells must be dynamic and communicate with one another.
Leukocytes of the immune system and neurons of the nervous system are well-accepted examples of cells that communicate and interact with differing cell populations. However, the formerly accepted notion that cells are static or possess only a single function is no longer tenable. Endothelial cells, once thought simply to line blood vessels, are now known both to respond to and secrete chemical signals (e.g., cytokines and growth factors) that enable communication with surrounding cells. In addition, endothelial cells express surface-bound leukocyte adhesion molecules that promote physical interaction with circulating blood cells. These adhesion molecules, which ultimately regulate leukocyte entrance into the underlying tissue, thereby participate in control of local inflammation. More recently, structural cells (such as fibroblasts and smooth muscle cells) have likewise been shown to receive and transmit chemical signals, as well as to interact with cells of the immune system via leukocyte adhesion molecules.
Hyaluronan also participates in the cellular reactions caused by environmental changes. In a previous contribution to this series,b a role was described for hyaluronan in the response of smooth muscle cells to physical vascular injury, which ultimately leads to restenosis. Hyaluronan can also participate in the reaction of a cell to microbial insult. Viral infection of smooth muscle cells, for example, elicits a hyaluronan response not seen in uninfected cells. This response (described below) facilitates smooth muscle cell interaction with leukocytes and, in this context, contributes to inflammation. 1
b See review by Wight in this series.
The large intestine is an organ whose principal function is to absorb water from the unusable portion of digested food as it passes along the bowel lumen. Hyaluronan is moderately abundant in colonic tissue and plays an important role in tissue osmosis. The colon is an elastic tube; the cellular organization of its internal mucosal layer is in many ways similar to that of the blood vessel (Fig.1A). The entire lumenal surface is covered with a monolayer of epithelial cells, beneath which is a network comprised of collagen fibers and smooth muscle cell bundles, referred to as the lamina propria. A thin layer of smooth muscle cells, the muscularis mucosae, lies directly under this network. Outside this inner mucosal layer is additional matrix and then a thicker layer of muscle cells, the muscularis propria. Under normal circumstances, the colonic mucosa contains a population of leukocytes, including T- and B-lymphocytes, plasma cells, histiocytes and mast cells, that arrive to the area via regularly distributed capillaries within the lamina propria. These leukocytes survey the mucosa and provide immune protection against the lumenal contents of the colon. Mucosal lymphocytes may reenter the blood stream, presumably via the lymphatic vessels located in close proximity to the muscularis mucosae, and are free to recirculate through blood and lymphoid organs until a specific antigenic challenge recalls them to an affected area.
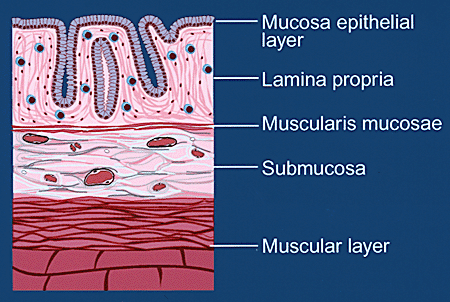
Fig. 1A Schematic diagram of tissue layers of normal colon.
Colon cancer and inflammatory bowel disease are two devastating and relatively common diseases of the large intestine. Colon cancer, a malignant disorder that affects an older population, typically metastasizes to other organ systems. Conversely, inflammatory bowel disease, a benign condition of younger individuals, is characterized by inflammation of the gut as well as of some extra-intestinal sites.
The metastatic potential of certain colon carcinomas correlates with the expression of the hyaluronan receptor CD44, which circumstantially links hyaluronan with tumor progression. CD44 is frequently overexpressed on many colon carcinomas, as are some of its isoforms.c The variant CD44v6 is most commonly expressed in advanced stages of tumor development and strongly correlates to colorectal tumor-related deaths.2 Recent evidence also suggests that stromal cells of the intestine synthesize hyaluronan in response to factors produced by adenocarcinomas.3
c See review by Knudson and Knudson in this series.
The origin of inflammatory bowel disease appears to be multifactorial; environmental and microbiological factors initiate and perpetuate an immune response in the intestine of genetically susceptible individuals. Some evidence suggests that hyaluronan may play a pivotal role in this inflammatory disorder. Increased hyaluronan levels have been measured in the intestine of patients with inflammatory bowel disease. Also, compared with other inflammatory disorders, greater levels of CD44 are observed throughout the inflamed colon tissue of patients with ulcerative colitis, a specific type of inflammatory bowel disease, and in particular increased amounts of CD44v6 and CD44v3 are expressed by the epithelial cells. Interestingly, the extra-intestinal manifestations of inflammatory bowel disease, which are exhibited in 30-50% of patients, occur in hyaluronan-rich tissues (e.g., eye, joint, and skin).
Immune phenomena linked to the etiopathogenesis of inflammatory bowel disease have also been extensively investigated, and findings have underscored the differences between the responses of normal and affected individuals. For example, it is well known that the distribution of specific cytokine-producing lymphocytes differs between normal persons and patients with Crohn's disease or ulcerative colitis. However, much less is known about how microbial agents affect the disease process. Speculations that viruses may be involved in the pathogenesis of inflammatory bowel disease have been advanced for some time because of the clinical association of respiratory virus infections with subsequent disease flares.
With inflammatory bowel disease, the mucosal immune cell population increases dramatically, and the infiltrate is predominantly comprised of mononuclear leukocytes (Fig. 1B). Furthermore, the muscularis mucosae cell layer thickens to nearly 300 times its normal depth secondary to smooth muscle cell hyperplasia and extracellular matrix deposition. These features suggest that interactions between recruited leukocytes and mesenchymal smooth muscle cells are important in the development and propagation of inflammatory bowel disease.
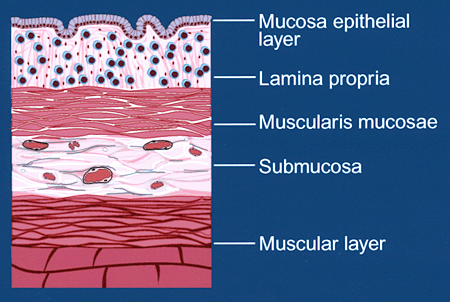
Fig. 1B Schematic diagram of tissue layers of colon with inflammatory bowel disease.
Leukocyte-produced, pro-inflammatory cytokines such as tumor necrosis factor (TNF)-alpha and interleukin-1 induce colon smooth muscle cell proliferation, which may contribute to hyperplasia. Additionally, these same cytokines, exemplified by TNF-alpha, cause smooth muscle cells to express increased levels of the leukocyte adhesion molecule known as vascular cell adhesion molecule-1 (VCAM-1) and to bind mononuclear leukocytes to their surface by means of this protein (Fig. 2). The act of adhesion to VCAM-1 by a subset of lymphocytes through their counter-receptor VLA-4 (very late antigen-4) can cause activation and production of 72-kD gelatinase by these lymphocytes, which may play a role in tissue remodeling.
Interestingly, viral agents can also increase VCAM-1 on a variety of cells, including colon smooth muscle cells. However, the mechanism of virus-induced leukocyte interaction with smooth muscle cells is very different than that induced by inflammatory cytokines (Fig. 2).
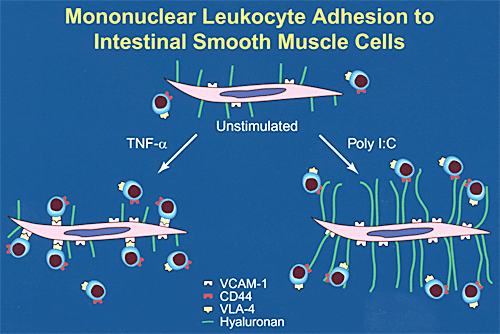
Fig. 2
Schematic diagram of leukocyte adhesion to inflammatory cytokine (TNF-alpha)-induced vs. virus (poly I:C)-induced leukocyte adhesion molecules on colon smooth muscle cells.
Cultured human colon smooth muscle cells are highly susceptible to infection by respiratory syncytial virus (RSV) and measles virus, two viruses circumstantially linked to inflammatory bowel disease. Although smooth muscle cells constitutively synthesize hyaluronan as part of their normal function, they respond to virus infection by elaborating cell surface-bound hyaluronan in a form that is highly adhesive for mononuclear leukocytes (i.e., T- and B-lymphocytes, monocytes, and mast cells). CD44, the major hyaluronan receptor, is the leukocyte ligand primarily responsible for this cell-cell interaction.
Phase contrast micrographs demonstrate the dramatic change in leukocyte adhesion to colon smooth muscle cells one day after infection with respiratory syncytial virus (Fig. 3). The leukocytes appear as bright spheres attached to the gray monolayer of smooth muscle cells. Poly I:C, a synthetic double-stranded RNA that mimics virus infection by activating many of the virus response pathways, but without killing the cells, similarly induces leukocyte adhesion to colon smooth muscle cells, as well as to smooth muscle cells isolated from vascular (mesenteric artery) and from airway (bronchus) tissue (Fig. 4).
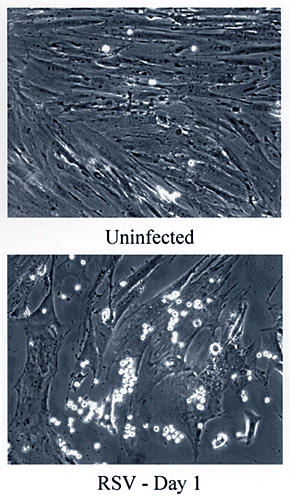
Fig. 3 Infection with respiratory syncytial virus (RSV) causes leukocyte adhesion to colon smooth muscle cells.
Confluent colon smooth muscle cells were treated with culture medium with or without RSV for 18 h at 37ºC. Unstimulated leukocytes were allowed to adhere for 1 h. The unbound leukocytes were washed away, and the cultures were observed by phase contrast microscopy (100x). Leukocytes appear as bright spheres on top of the smooth muscle cells, which are attached to the culture plate.
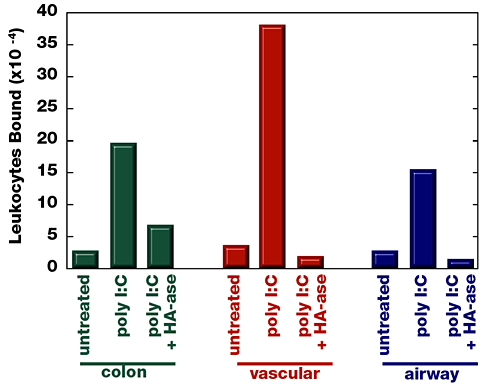
Fig. 4
Poly I:C treatment causes increased leukocyte adhesion to colon, vascular, and airway smooth muscle cells that is abrogated by hyaluronidase digestion. Confluent smooth muscle cells were treated overnight with medium with or without the viral mimic, poly I:C. Testicular hyaluronidase was added to indicated culture wells, followed by incubation for 10 min at 37ºC. The binding of radioactively labeled leukocytes is indicated.
The majority of leukocyte adhesion is mediated by hyaluronan on the virus-treated or poly I:C-treated smooth muscle cell surface. This is demonstrated in Figure 4, which shows that digestion with hyaluronidase abrogates most of the leukocyte adhesion. Residual leukocyte binding is mediated by VCAM-1 on the smooth muscle cells and can be blocked by a specific anti-VCAM-1 monoclonal antibody, as described above. Patient variation accounts for the relative contribution by each adhesion molecule to total mononuclear leukocyte adhesion.
As mentioned above, virus infection or poly I:C treatment results in increased smooth muscle cell surface expression of VCAM-1 (usually at higher levels than those induced by TNF-alpha) and hyaluronan, but only hyaluronan mediates significant levels of leukocyte adhesion. When hyaluronan is enzymatically removed from the cell surface, VCAM-1 is fully capable of binding leukocytes. These findings suggest another interesting function for hyaluronan, that of masking other adhesion molecules (Fig. 2). Hyaluronan-mediated leukocyte adhesion to cultured smooth muscle cells has a distinctly different appearance from leukocyte adhesion via VCAM-1. The phase-contrast micrographs (Fig. 5) depict leukocytes binding to poly I:C treated smooth muscle cells via hyaluronan. Chain-like formations of leukocytes that can span several cell lengths are obvious. Removal of hyaluronan, by hyaluronidase treatment, results in the appearance of singly bound leukocytes, typical of VCAM-1-mediated leukocyte adhesion.

Fig. 5 Phase contrast micrographs of poly I:C-induced leukocyte adhesion to colon smooth muscle cells.
Leukocytes binding to poly I:C treated smooth muscle cells frequently appear to line up in chains or clumps. Hyaluronidase treatment prior to leukocyte adhesion exposes another molecule, VCAM-1, which results in the distinctly different pattern of adhesion shown in the rightmost panel.
Cultured colon smooth muscle cells retain proportionally little (~20% after 15 h culture) of the hyaluronan they synthesize under basal conditions. In response to poly I:C general hyaluronan synthesis increases, and the proportion that is cell associated also increases (~35%). The hyaluronan synthesized by poly I:C-treated smooth muscle cells is incorporated into two distinct structural forms that are not seen with untreated cells. Confocal microscopic visualization of colon smooth muscle cells co-stained for hyaluronan (fluorescein) and CD44 (Texas red) reveals that poly I:C induces production of pericellular hyaluronan coats, as well as strand-like hyaluronan structures that span several cell lengths (Fig. 6A). The smooth muscle cells also express abundant amounts of CD44 on their surfaces, but this expression is not modulated by poly I:C.
Non-activated leukocytes (identified by their blue nuclear stain) similarly express CD44 on their cell surface (Texas red) but do not bind to hyaluronan on untreated smooth muscle cells. However, they do bind avidly to the virus-induced hyaluronan of the muscle cells via CD44 (Fig. 6B), but this interaction appears selective, as the CD44 on the leukocytes binds only to the strand-like hyaluronan structures, not to the pericellular hyaluronan coats.
The factors that contribute to the organization of hyaluronan into these macromolecular structures that favor the binding of non-activated leukocytes are yet to be defined, but likely include several of the hyaluronan-binding proteins. One of the potential candidates is the serum protein inter-alpha-trypsin inhibitor (I-alpha-I or ITI), which has been described as a stabilizer of hyaluronan pericellular coats on a variety of cells, including smooth muscle cells. Similarly, hyaladherin molecules such as aggrecan, versican, hyaluronectin, or the protein produced by TNF-stimulated gene-6 (TSG-6) may play a role in forming hyaluronan into these strand-like structures and possibly interact with leukocyte CD44.
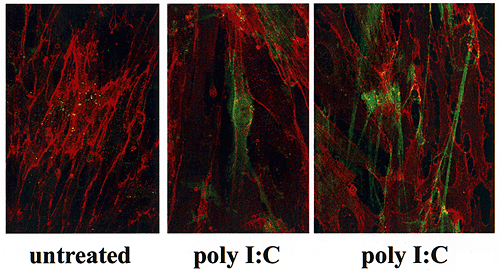
Fig. 6A Confocal micrographs of poly I:C-induced hyaluronan and CD44 on colon smooth muscle cells.
Colon smooth muscle cells were dual-stained for hyaluronan and CD44. The hyaluronan-binding protein was used with a secondary flurorescein indicator, which displays the hyaluronan in green. To detect CD44, the monoclonal antibody to CD44 (A3D8) was used in conjunction with a Texas red-labeled secondary antibody.
(A) Untreated smooth muscle cells have little hyaluronan (green) on their cell surface. After poly I:C treatment, cocoon-like pericellular hyaluronan matrices (middle panel) are obvious in the culture, as well as strand-like hyaluronan structures (right panel) that span several cells.
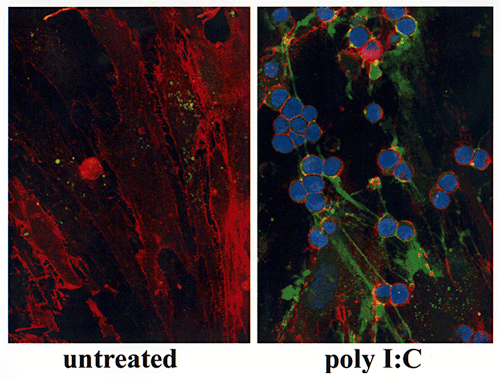
Fig. 6B
(B) Leukocyte (blue nuclear stain) adhesion is mediated by the strand-like structures. CD44 on the leukocyte surface (red) mediates their adhesion to hyaluronan.
Much is known about the binding of CD44, the major hyaluronan receptor, to its ligand.c Recent findings have certainly underscored the potential importance of the CD44 receptor/hyaluronan interaction to inflammation. For example, specific ligation of CD44 to hyaluronan activates peripheral blood T-cells, causing increased IL-2 levels, and activates monocytes to release higher levels of the pro-inflammatory cytokines IL-1 and TNF-alpha. Similarly, activation of T-cell-associated CD44 4 is required for leukocyte extravasation into inflammatory sites, and CD44 is critical to the secondary leukocyte recruitment observed with inflammation of the central nervous system and experimental encephalomyelitis.5 In other tissues, binding of hyaluronan to macrophages has been shown to upregulate chemokine expression (i.e., RANTES, MIP-1a, and MIP-1b), as well as the production of IL-12.6 Since viral agent-induced hyaluronan on intestinal smooth muscle cells binds non-activated leukocytes through CD44, subsequent activation of the leukocytes likely follows, thereby exacerbating the inflammatory response.
The reason that colon smooth muscle cells under virus attack elaborate hyaluronan in strands that are adhesive for non-activated leukocytes is poorly understood. Current literature suggests that CD44 activation is obligatory for its binding to hyaluronan, yet this requisite appears unnecessary with the virus-induced mechanism of adhesion. Unlike binding to traditional leukocyte adhesion molecules (e.g., VCAM-1), leukocyte binding to strand-like hyaluronan structures permits different types of mononuclear leukocytes to interact with each other and further facilitate an immune response. Hyaluronan may create a microenvironment in which closely proximate bound leukocytes can communicate directly by surface contact or indirectly through the production of soluble cytokine concentrates. The novel hyaluronan response witnessed with virus infection provides compelling impetus for further investigative studies to better understand the pathogenesis of intestinal diseases and chronic inflammatory disorders.



