Hayato Ota
Research Assistant Lecturer, Glycan and Life Systems Integration Center (GaLSIC), Soka University, Japan.
He received his Ph.D. in Engineering from the Department of Bioinformatics, Graduate School of Engineering, Soka University, in 2023. He served as an Assistant Lecturer in the Faculty of Science and Engineering at Soka University from September 2023 to September 2025. Since October 2025, he has been a Research Assistant Lecturer at GaLSIC, Soka University.

Kazuo Yamamoto Ph.D.
Visiting Professor, Institute for Human Life Science, Ochanomizu University
He received his Ph.D. from the Graduate School of Pharmaceutical Sciences, The University of Tokyo, in 1984. After serving as a Researcher at the Tokyo Metropolitan Institute of Medical Science, a Research Associate at the Faculty of Pharmaceutical Sciences, The University of Tokyo, and an Associate Professor at the Graduate School of Pharmaceutical Sciences, The University of Tokyo, he was appointed Professor at the Graduate School of Frontier Sciences, The University of Tokyo, where he served from 1999 to 2022. He became Professor Emeritus of The University of Tokyo in 2022. After serving as a Project Professor at the Graduate School of Medicine, Chiba University, he currently serves as a Visiting Professor at Ochanomizu University.

Shoko Nishihara Ph.D.
Director and Distinguished Professor, Glycan and Life Systems Integration Center (GaLSIC), Soka University
She received her Ph.D. from The University of Tokyo in 1982. After working at Keio University, The Jikei University School of Medicine, The University of North Carolina at Chapel Hill, and the Mitsubishi Kasei Institute of Life Sciences, she joined Soka University in 1991. She currently serves as President of the Japan Consortium for Glycobiology and Glycotechnology (JCGG) and as an Editorial Board Member of Glycoconjugate Journal. Her research focuses on the conserved functions of glycans across species, particularly in mammalian pluripotent stem cells and Drosophila. She is also involved in collaborative studies aimed at developing screening methods and conducting functional analyses for undiagnosed and rare glycan-related diseases.
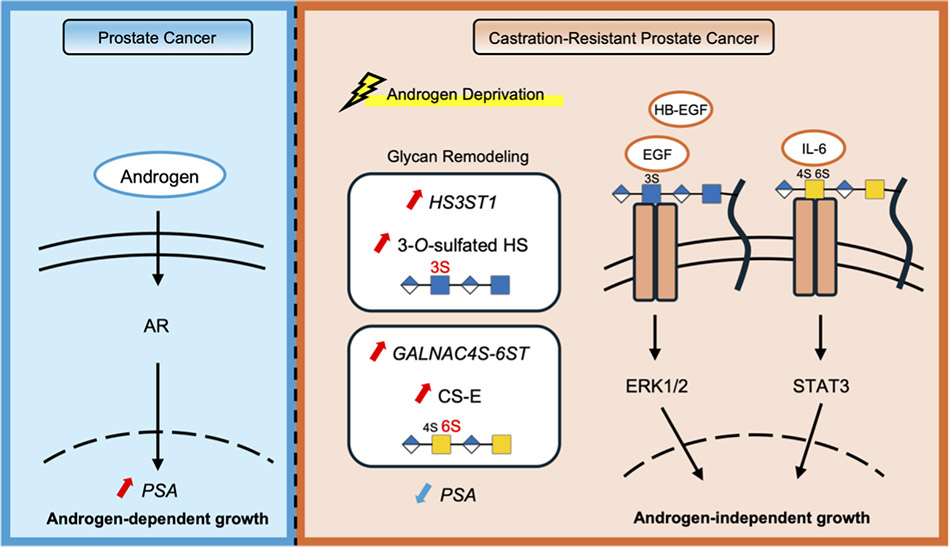
Prostate cancer grows in an androgen receptor (AR)-dependent manner; however, some cases progress to castration-resistant prostate cancer (CRPC) after androgen deprivation therapy. One of the molecular mechanisms underlying this transition is the activation of AR-independent “bypass signaling” pathways. Although several signaling pathways have been implicated, the mechanisms that regulate these signaling pathways remain incompletely understood. In this article, we focus on cell surface glycans, particularly sulfated glycosaminoglycans (GAGs), and summarize two independent mechanisms involved in the acquisition of castration resistance: activation of epidermal growth factor receptor (EGFR) signaling mediated by 3-O-sulfated heparan sulfate (3-OS HS) and enhancement of interleukin (IL)-6/signal transducer and activator of transcription (STAT)3 signaling mediated by chondroitin sulfate E (CS-E). These findings highlight the potential of GAGs as novel therapeutic targets in CRPC.
Prostate cancer is one of the most common malignancies in men and typically grows in an androgen receptor (AR)-dependent manner. Androgen deprivation therapy is therefore widely used as the primary treatment. However, in a subset of patients, tumors acquire resistance to therapy and progress to castration-resistant prostate cancer (CRPC)1. In CRPC, not only reactivation of AR signaling but also alternative proliferative pathways independent of AR, often referred to as “bypass signaling,” are thought to play important roles in tumor progression. Nevertheless, the integrated mechanisms that regulate these signaling pathways remain incompletely understood2-4.
In recent years, studies in cancer biology and clinical diagnostics have revealed that not only protein expression levels but also the glycan structures attached to proteins reflect disease states and tumor malignancy. In prostate cancer, efforts have been made to improve diagnostic accuracy by analyzing the glycan structures of prostate-specific antigen (PSA), a widely used diagnostic biomarker5. Meanwhile, glycans are not merely molecular markers but also function as regulatory molecules that modulate receptor–ligand interactions at the cell surface and thereby control intracellular signaling pathways6,7. Sulfated glycosaminoglycans (GAGs), such as heparan sulfate and chondroitin sulfate, mediate the binding of diverse growth factors and cytokines and play crucial roles in regulating cancer cell proliferation, survival, and responses to the microenvironment8.
In this article, we focus on the remodeling of glycan structures that occurs as prostate cancer cells adapt to hormone-deprived conditions. We summarize two glycan-dependent mechanisms underlying the acquisition of castration resistance: activation of epidermal growth factor receptor (EGFR) signaling mediated by 3-O-sulfated heparan sulfate (3-OS HS) and regulation of interleukin (IL)-6/signal transducer and activator of transcription (STAT)3 signaling mediated by chondroitin sulfate E (CS-E). Through these findings, we discuss the possibility that glycans provide a molecular basis that determines signaling selectivity in CRPC.
Cells receive information from their surrounding environment through receptors located on the plasma membrane, and these extracellular signals thereby regulate diverse cellular processes such as proliferation, differentiation, and survival. Intracellular signal transduction is typically initiated when ligands, including growth factors and cytokines, bind to their corresponding receptors. However, these interactions are not always mediated solely by receptors and ligands. Glycans present on the cell surface can modulate the formation of receptor complexes and the efficiency of ligand binding, thereby functioning as “co-receptors” in signal transduction6,7.
Among various glycans, glycosaminoglycans (GAGs) are linear polysaccharides abundantly present on the cell surface and within the extracellular matrix. GAGs are composed of repeating disaccharide units consisting of an amino sugar, such as N-acetylglucosamine or N-acetylgalactosamine, and a uronic acid, such as glucuronic acid or iduronic acid. Based on their structural features, GAGs are classified into several groups, including heparan sulfate (HS), chondroitin sulfate (CS), dermatan sulfate (DS), and keratan sulfate (KS).
A key functional property of GAGs lies not in the glycan backbone itself but in the pattern of sulfation, namely the positions and number of sulfate groups attached to the polysaccharide chain. Differences in sulfation patterns determine ligand-binding specificity, resulting in distinct receptor activation and downstream signaling pathways even within the same class of GAGs. In this way, sulfation patterns function as a molecular basis that enables cells to selectively interpret diverse environmental signals. Thus, GAGs should not be regarded merely as structural components of the cell surface but rather as active regulatory molecules that modulate the efficiency and selectivity of extracellular signal reception.
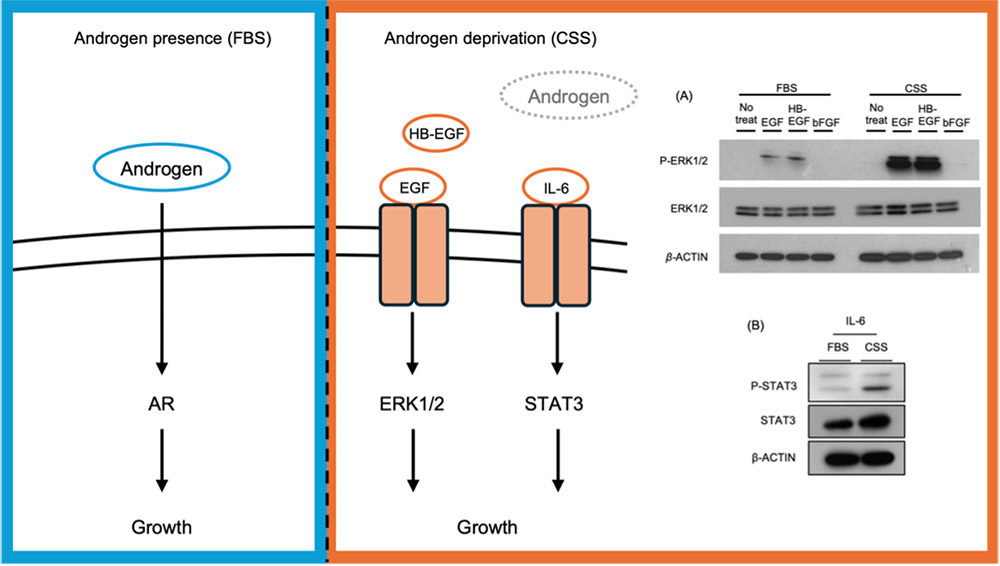
The CRPC cell line C4-2 can proliferate under both hormone-replete conditions using fetal bovine serum (FBS) and hormone-deprived conditions using charcoal-stripped serum (CSS)9-11. Analysis of signaling changes under these culture conditions revealed that, in CSS-containing medium, both epidermal growth factor (EGF) or heparin binding (HB)-EGF/EGFR signaling and IL-6/STAT3 signaling are enhanced in place of AR signaling (Fig. 1). Although these pathways have been recognized as androgen-independent bypass signaling mechanisms, the detailed molecular basis underlying their activation remains unclear.
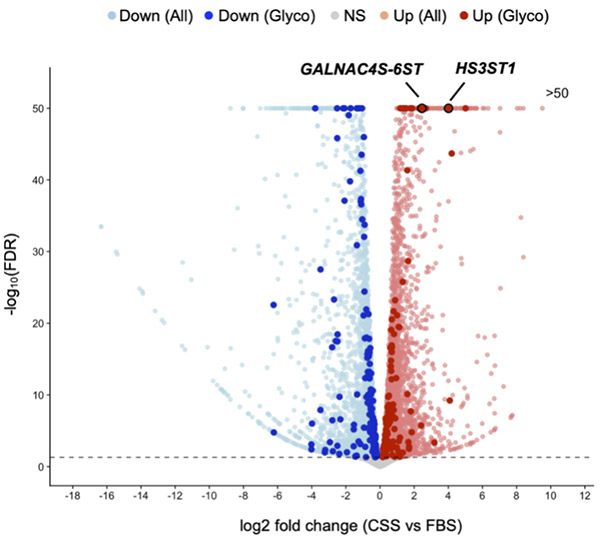
Recent studies have suggested that alterations in the sulfation status of GAGs occur in response to hormone deprivation and may contribute to AR-independent proliferation in CRPC10,11. RNA-seq analysis comparing gene expression in C4-2 cells cultured in FBS and CSS-containing medium revealed extensive transcriptional changes (Fig. 2). Among glycan-related genes, 131 genes were significantly upregulated, and 151 genes were significantly downregulated under hormone-deprived conditions, indicating dynamic remodeling of glycan-related gene expression. Notably, the expression of HS3ST1, a heparan sulfate 3-O-sulfotransferase, and GALNAC4S-6ST, an enzyme responsible for the synthesis of CS-E, was markedly increased.
These findings suggest that hormone deprivation not only alters transcriptional networks but also remodels the composition of cell surface glycans. In CRPC cells, regulation of glycan-related gene expression may therefore generate a cell surface environment that facilitates the activation of AR-independent signaling pathways.
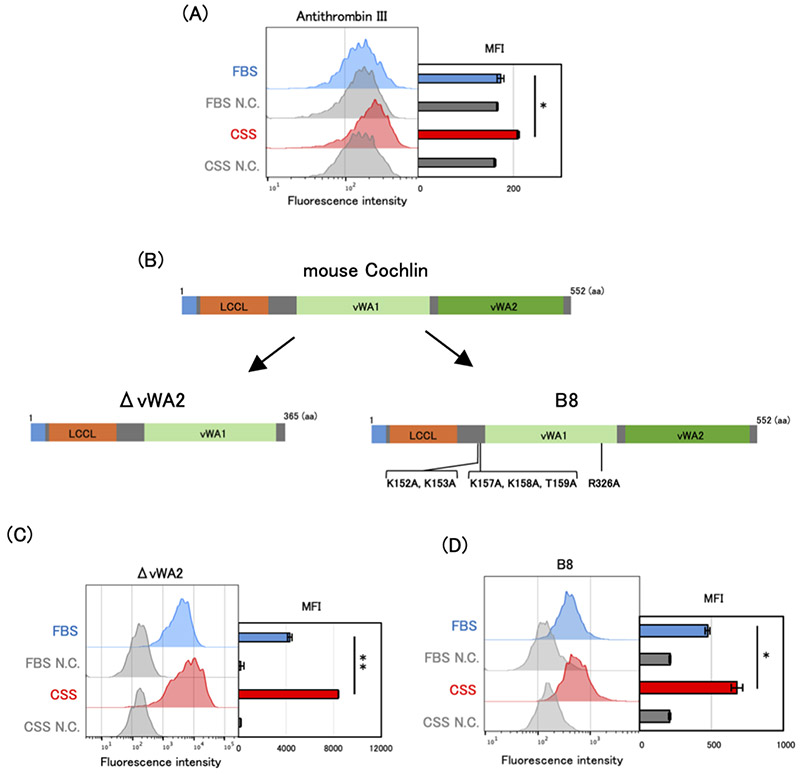
Sulfated glycosaminoglycans (GAGs) represent a class of molecules whose functional analysis has long been limited due to their structural diversity and analytical complexity. Selectively identifying specific sulfation patterns on the cell surface has been technically challenging, and conventional antibodies or lectins often lack sufficient specificity. In recent years, engineered variants of Cochlin, a lectin that binds sulfated glycans, have been developed to selectively recognize sulfated structures11-13. These engineered Cochlin proteins exhibit high affinity for highly sulfated glycan structures and provide a means to directly visualize sulfated glycans on the cell surface using techniques such as flow cytometry and tissue staining.
Mouse Cochlin is a lectin that primarily recognizes heparin and CS-E and consists of three domains: an LCCL domain, a vWA1 domain, and a vWA2 domain. An engineered Cochlin lacking the vWA2 domain (ΔvWA2 Cochlin) recognizes a broader range of sulfated GAGs than wild-type Cochlin. In contrast, B8 Cochlin, which carries six amino acid substitutions (K152A, K153A, K157A, K158A, T159A, and R236A), is an engineered variant with enhanced specificity and affinity for heparin and CS-E compared with the wild-type protein (Fig. 3).
In the studies described in this article on 3-OS HS and CS-E, these Cochlin-based glycan recognition tools played a critical role10,11. Specifically, the cell surface of C4-2 cells cultured in FBS- or CSS-containing medium was stained with engineered Cochlin and analyzed by flow cytometry. Using ΔvWA2 Cochlin, which recognizes sulfated GAGs including 3-OS HS, staining intensity increased under hormone-deprived conditions. Moreover, knockdown of HS3ST1 reduced the staining signal, and a similar trend was observed using Antithrombin III, which specifically recognizes 3-OS HS (Fig. 3A–C). These results indicate that sulfated GAGs containing 3-OS HS increase under hormone-deprived conditions.
Similarly, increased staining intensity under CSS-containing culture conditions was also observed with B8 Cochlin, which recognizes CS-E, indicating that cell surface expression of CS-E is elevated under hormone-deprived conditions (Fig. 3D). The development of such molecular tools capable of recognizing specific glycan structures has become a major driving force in advancing glycobiology research.
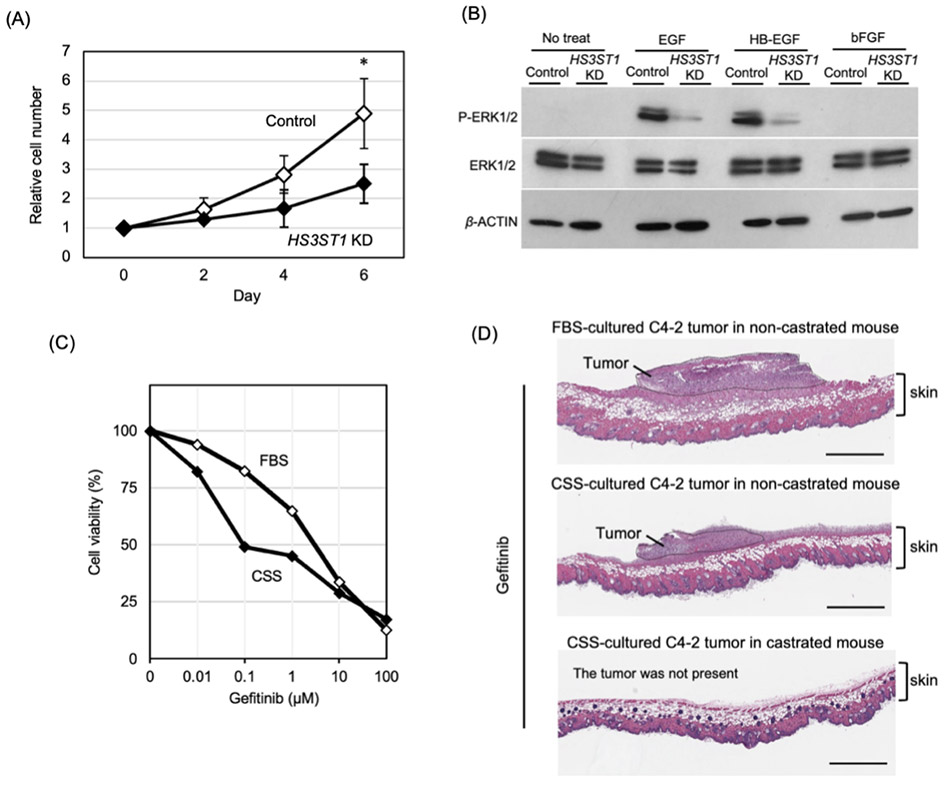
As described above, increased expression of the heparan sulfate 3-O-sulfotransferase HS3ST1 in CRPC leads to an elevation of 3-O-sulfated heparan sulfate (3-OS HS) structures. 3-O-sulfation transferred by the HS3ST family has been reported to regulate intracellular signaling pathways14-16. To investigate whether the increase in 3-OS HS mediated by HS3ST1 contributes to AR-independent proliferation in CRPC, knockdown of HS3ST1 was performed10. In HS3ST1 knocked down C4-2 cells cultured in CSS-containing medium, cell proliferation was reduced and EGFR signaling activity was markedly suppressed (Fig. 4A,B). These findings indicate that 3-OS HS promotes cell proliferation in CRPC through activation of EGFR signaling.
To further evaluate the functional importance of EGFR signaling, the growth-inhibitory effects of the EGFR inhibitor gefitinib were compared under hormone-replete and hormone-deprived conditions. C4-2 cells cultured in CSS-containing medium showed significantly greater sensitivity to gefitinib than those cultured in FBS-containing medium (IC50: 3.67 μM in FBS vs. 0.33 μM in CSS) (Fig. 4C). C4-2 cells cultured in medium containing FBS or CSS were subcutaneously transplanted into intact or castrated mice and treated with gefitinib. Greater tumor growth inhibition was observed in tumors formed under more androgen-deprived conditions (Fig. 4D). These results suggest that CRPC cells become more dependent on EGFR signaling under hormone-deprived conditions.
At the molecular level, 3-OS HS may facilitate the interaction of EGFR ligands such as EGF and HB-EGF with their receptor, thereby enhancing EGFR signaling. Importantly, this activation appears to result not from changes in ligand or receptor expression levels but from alterations in the sulfation patterns of cell surface glycans. Taken together, these results suggest that 3-OS HS promotes EGFR signaling under hormone-deprived conditions, enabling CRPC cells to shift their signaling dependency from AR-mediated proliferation to EGFR-dependent growth. Thus, 3-O-sulfation of HS represents a key molecular mechanism supporting AR-independent proliferation in CRPC.
When C4-2 cells are cultured under hormone-deprived conditions using CSS, the expression of GALNAC4S-6ST, a key enzyme responsible for the synthesis of CS-E, is increased, leading to elevated cell surface expression of CS-E. CS-E has been reported to regulate intracellular signaling pathways in various cell types17-21.
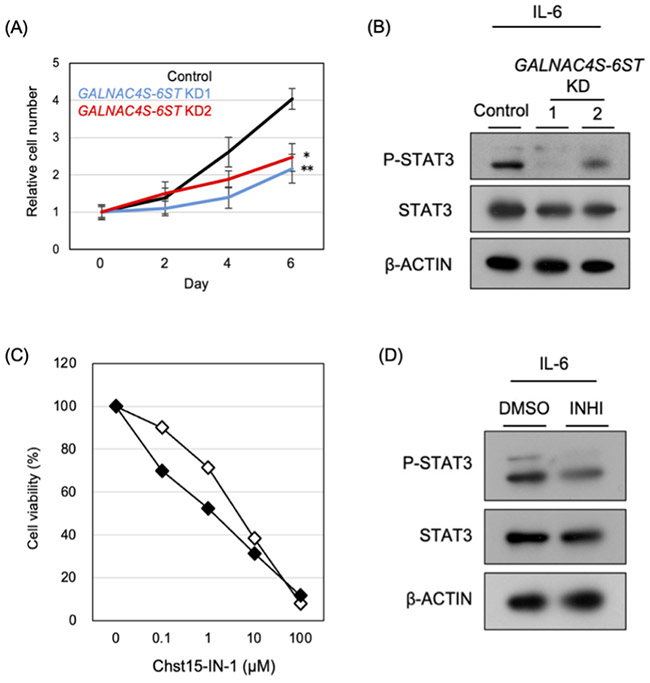
To investigate the role of CS-E in the acquisition of castration resistance, the expression of GALNAC4S-6ST was suppressed in C4-2 cells11. Knockdown of GALNAC4S-6ST resulted in reduced cell proliferation (Fig. 5A). At the same time, activation of IL-6/STAT3 signaling was markedly suppressed (Fig. 5B). In contrast, other signaling pathways such as EGFR and Wnt were not significantly affected, suggesting that CS-E selectively regulates IL-6/STAT3 signaling in CRPC. Mechanistically, CS-E appears to enhance the binding of IL-6 to the cell surface and stabilize the formation of the IL-6 receptor complex, thereby promoting STAT3 phosphorylation. In addition, previous studies have reported that CS-A, a precursor structure of CS-E, is involved in the progression of CRPC22,23. These findings suggest that increased CS-A levels may lead to enhanced CS-E production, thereby contributing to the activation of CS-E–mediated proliferative signaling.
The functional significance of CS-E was further examined using the GALNAC4S-6ST inhibitor Chst15-IN-1. Treatment of C4-2 cells cultured in FBS- or CSS-containing medium revealed the stronger growth-inhibitory effect under CSS culture conditions (IC50: 4.46 μM in FBS vs. 1.39 μM in CSS) (Fig. 5C). Consistent with the knockdown experiments, this inhibitor also reduced the level of STAT3 phosphorylation (Fig. 5D). Taken together, these results suggest that CRPC cells become increasingly dependent on the CS-E/IL-6/STAT3 signaling axis to maintain proliferation. Thus, activation of CS-E–dependent IL-6/STAT3 signaling represents an important molecular mechanism promoting AR-independent growth in CRPC.
The two pathways described in this article—EGFR signaling mediated by 3-OS HS and IL-6/STAT3 signaling mediated by CS-E—represent AR-independent bypass signaling mechanisms that are activated in CRPC. Although these pathways utilize distinct receptors and downstream signaling cascades, they share a common feature in that their signaling efficiency is regulated by changes in the sulfation patterns of cell surface glycans. Importantly, activation of these signaling pathways is not driven by increased expression of ligands or receptors but rather by alterations in glycan structures on the cell surface. In response to the environmental change caused by hormone deprivation, prostate cancer cells appear to remodel glycan biosynthesis and sulfation pathways, thereby creating a cell surface environment that efficiently captures growth factors and cytokines. As a result, cancer cells can maintain alternative proliferative and survival signaling even when AR signaling is suppressed. 3-OS HS and CS-E promote distinct signaling pathways but may be part of a common molecular strategy that enables cancer cells to adapt to environmental changes.
Another important aspect is that these two glycan-dependent signaling pathways are not mutually exclusive but may function in parallel within the same cell. Diversification of glycan structures may therefore generate multilayered signaling redundancy in CRPC, which could contribute to the difficulty of treating CRPC. Taken together, these findings indicate that glycans function as active regulatory determinants of signaling selectivity in CRPC. Understanding signaling networks from the perspective of glycan structural remodeling may provide a more comprehensive view of CRPC pathogenesis and new opportunities for therapeutic development.