Yusuke Mimura
Department of Clinical Research, National Hospital Organization Yamaguchi Ube Medical Center
Yusuke Mimura did his M.D. and Ph.D. in medical sciences at Yamaguchi University. He did postdoctoral works at The University of Birmingham, The Glycobiology Institute at The University of Oxford, The University of Southampton, and later served as a senior research fellow in glycobiology at NIBRT. He then moved to National Hospital Organization Yamaguchi Ube Medical Center as the head of the Department of Clinical Research. His research interests include the development of novel IgG antibody therapeutics through glycoengineering.
1. Abstract
IgG activates FcγRs and the complement system while intravenous immunoglobulin (IVIG) consisting of plasma IgG exerts anti-inflammatory effects in the treatment of autoimmune diseases. Although IVIG has been used for >40 years as a treatment option for certain autoimmune disorders, its mechanism of action (MOA) remains unsolved. Recently developed chemo-enzymatic glycoengineering approach allows for remodeling of the Fc glycans of polyclonal IgG. We prepared fucosylated or nonfucosylated plasma IgG glycoforms having 2 sialic acids, 2 galactose, or 0 galactose at the nonreducing ends of the Fc glycans to investigate their anti-inflammatory activity. We have demonstrated that the galactosylated, nonfucosylated [(G2)2] glycoform has the highest affinity for FcγRIIIa and potency to inhibit antibody-dependent cellular cytotoxicity (ADCC) activity of immune cells. As the (G2)2 glycoform is a component of IVIG, glycoengineered IVIG consisting of the (G2)2 glycoform alone may be a potential next-generation antibody drug. This review outlines our current understanding of the role of IgG-Fc glycan in MOA of IVIG.
2. The influence of the glycan structure of IgG on Fc effector functions
IVIG is a therapeutic preparation of normal polyclonal IgG derived from pooled plasma of thousands of healthy donors. IVIG is used at a high-dose in the treatment of autoimmune/inflammatory disorders, including immune thrombocytopenia (ITP), Kawasaki Disease and Guillain-Barré syndrome1. Although the anti-inflammatory activity of IVIG is shown to reside in the Fc portion of IgG from a clinical study on treatment of ITP with the Fc fragments, its MOA remains unclear2. Because Fc glycosylation is required for the anti-inflammatory activity of IVIG3, we focused on the influence of the Fc glycan structures on the anti-inflammatory activity of IVIG4.
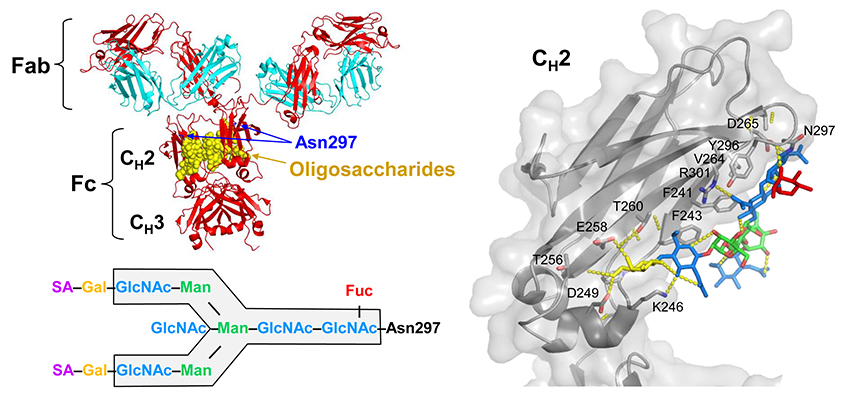
Fig. 1. Structures of IgG and the Fc glycan attached at Asn297 residue of each CH2 domain
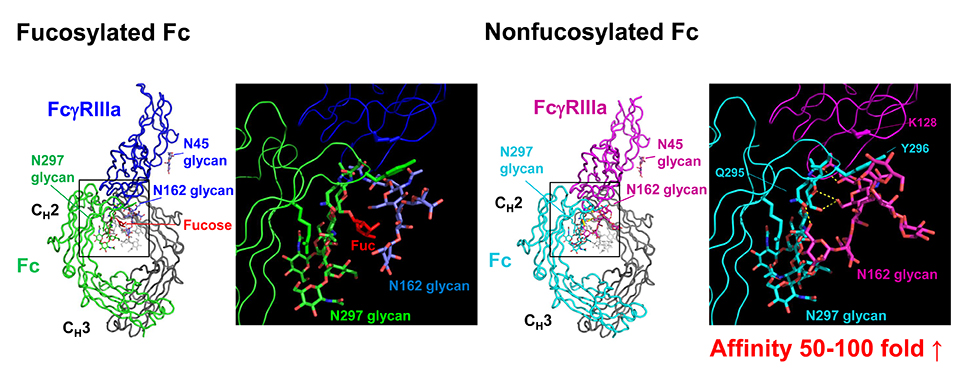
The biantennary complex-type glycans attached at Asn297 residues of IgG-Fc (Fig. 1) are essential for optimal expression of biological activities mediated through Fcγ receptors (FcγRs) and the C1q component of complement (antigen phagocytosis, superoxide release, ADCC, complement-dependent cytotoxicity, etc.). Removal of the Fc glycans can abrogate or severely compromise the Fc effector functions5,6. IgG-Fc glycans are heterogeneous due to variable addition and processing of outer-arm sugar residues (sialic acid, galactose, and bisecting GlcNAc) and fucose onto the core biantennary heptasaccharide (GlcNAc2Man3GlcNAc2) (Fig. 1). The differentially glycosylated species (glycoforms) are known to express unique biological activities7-9. In particular, defucosylation of the Fc glycans increases affinity for FcγRIIIa 50 – 100-fold and enhances ADCC (Fig. 2) 10-13.
Fig. 2. Crystal structures of the complexes of fucosylated Fc (Left) and nonfucosylated Fc (Right) with FcγRIIIa12
3. The Interactions between IgG and FcγRs
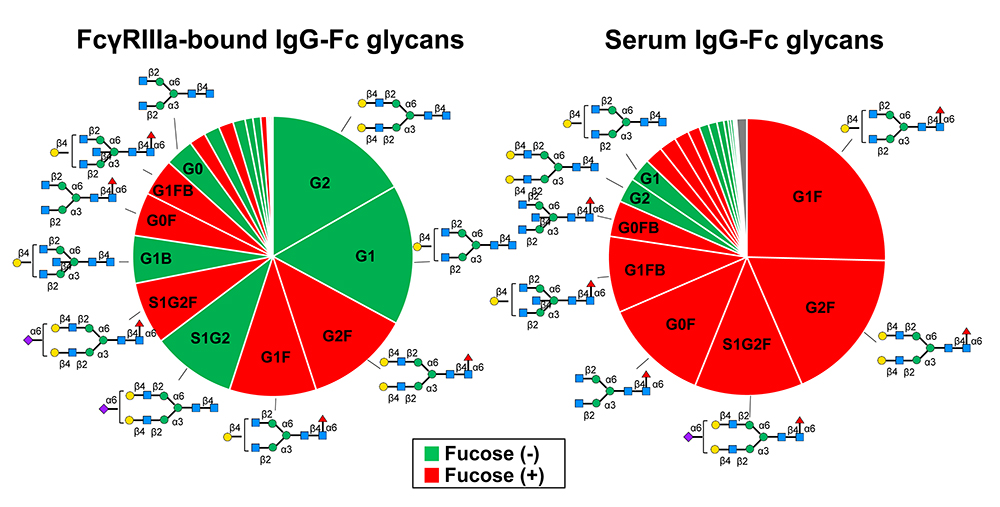
The family of human FcγRs that bind human IgG-Fc can be classified into five activating receptors (FcγRI, FcγRIIa, FcγRIIc, FcγRIIIa, FcγRIIIb) and one inhibitory receptor (FcγRIIb), and individual receptors are constitutively expressed on most immune cells in a cell-type specific manner14. FcγRs are also classified into high-affinity FcγR (FcγRI, Ka >108 M-1) and low-affinity FcγRs (FcγRII and FcγRIII, Ka <107 M-1). High-affinity FcγRI on circulating immune cells binds monomeric IgG15. On the other hand, low-affinity FcγRII and FcγRIII bind polymeric IgG, i.e., immune complexes readily and bind monomeric IgG to some extent in blood4,16. It has recently been reported that FcγRIIIa molecules on circulating NK cells preferentially bind nonfucosylated IgG17 (Fig. 3). Due to the asymmetry of the IgG-Fc/FcγRIIIa interaction non-fucosylation of one heavy chain would be sufficient for tight binding. As the majority of the Fc glycans released from IgG1 that had bound FcγRIIIa were nonfucosylated (Fig. 3, left), it is likely that nearly all FcγRIIIa molecules on NK cells are saturated by IgG lacking fucose on the condition that all the nonfucosylated H chains are paired with fucosylated H chains.
Fig. 3. Difference in the fucosylation levels of the Fc glycans between IgG bound to FcγRIIIa on NK cells (Left) and serum IgG of the same donors (Right)17
The enhanced affinity of nonfucosylated IgG for FcγRIIIa is attributed to the carbohydrate–carbohydrate interactions as well as the protein–protein interactions between IgG-Fc and FcγRIIIa (Fig. 2)12,13. Therefore, it is presumed that, due to the high affinity for FcγRIIIa, nonfucosylated IgG in serum can compete with immune complexes for FcγRIIIa. Importantly, the binding of monomeric IgG to FcγR in a 1:1 stoichiometry does not trigger activation of immune cells8,18. So, we hypothesized that high-affinity binding of serum nonfucosylated IgG to FcγRIIIa could inhibit ADCC activity of NK cells.
4. Anti-inflammatory effect of IVIG glycoforms
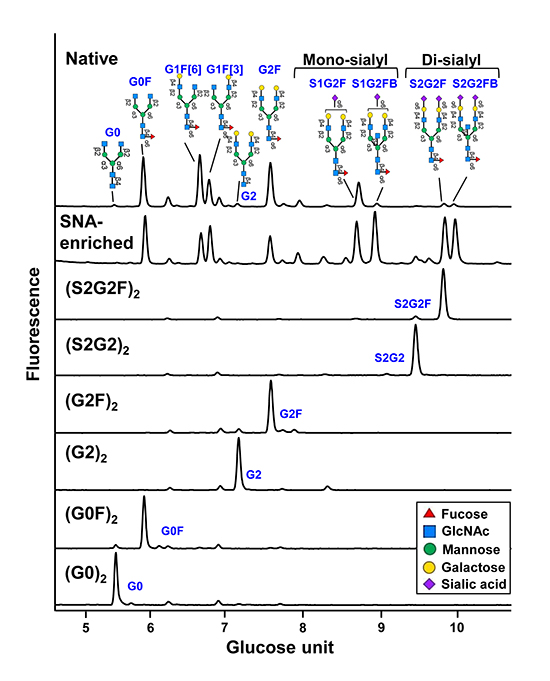
First, we remodeled normal plasma IgG glycans to prepare fucosylated [(S2G2F)2, (G2F)2 and (G0F)2] or nonfucosylated glycoforms [(S2G2)2, (G2)2 and (G0)2] of IgG bearing sialylated, galactosylated or nongalactosylated glycans (Fig. 4)19,20.
Fig. 4. HILIC analysis of the Fc glycans of native plasma IgG, SNA-enriched IgG, and glycoengineered IgG glycoforms Note that the glycan profiles of glycoengineered IgG glycoforms exhibit single Fc glycan peaks, in contrast to the heterogeneous glycan profile of SNA-enriched IgG.
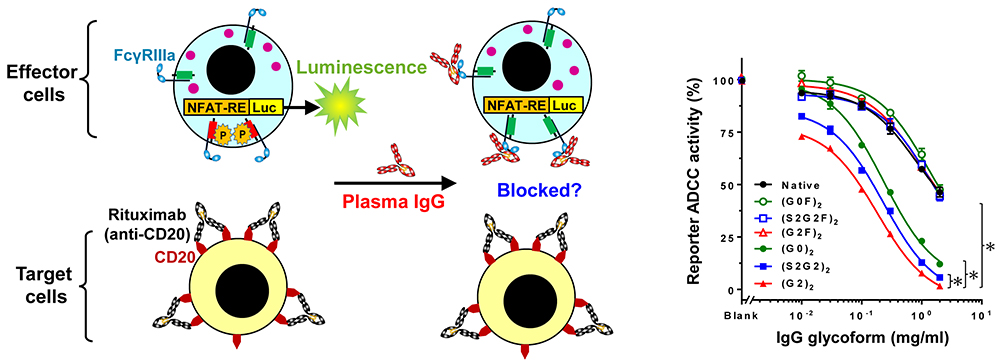
Engagement of FcγRIIIa on an immune cell by an IgG-opsonized target cell can induce ADCC. We investigated whether IgG glycoforms could compete with opsonizing CD20 antibody for FcγRIIIa and inhibit ADCC (Fig. 5, Left).
Fig. 5. Inhibition of ADCC by plasma IgG glycoforms ADCC reporter assay (Left). Titration of IgG glycoforms for their ADCC inhibition (Right)
As a result, ADCC was inhibited with IgG glycoforms in descending order of (G2)2>(S2)2>(G0)2>Native IgG (Fig. 5, Right), which indicates that (G2)2 has 20-fold higher capability to inhibit ADCC4.
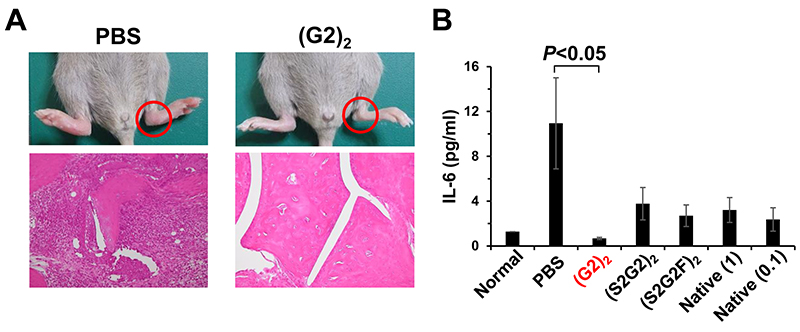
Furthermore, a murine collagen antibody-induced arthritis model was used to investigate whether the IVIG glycoforms exert the anti-inflammatory activity in vivo. Interestingly, (G2)2 IVIG reduced inflammation more effectively than the 10-fold higher dose of native IVIG, in contrast to sialylated IVIG glycoforms4 (Fig. 6).
Fig. 6. Anti-inflammatory effect of homogeneous IVIG glycoforms Anti-collagen antibodies were injected intravenously to DBA/1J mice on day 0. The IVIG glycoforms(native、(S2G2)2、(S2G2F)2、(G2)2)at 0.1 g/kg, positive control native IVIG at 1 g/kg, and PBS were administered on day 3. (A) Histology of the hind paw of a mouse treated with either PBS or (G2)2 IVIG on day 8. (H&E staining, magnification 100 x). (B) Serum IL-6 levels on day 8 (one-way ANOVA with Tukey's multiple-comparison test).
5. Importance of FcγRIIIa blockade in the treatment of autoimmune diseases
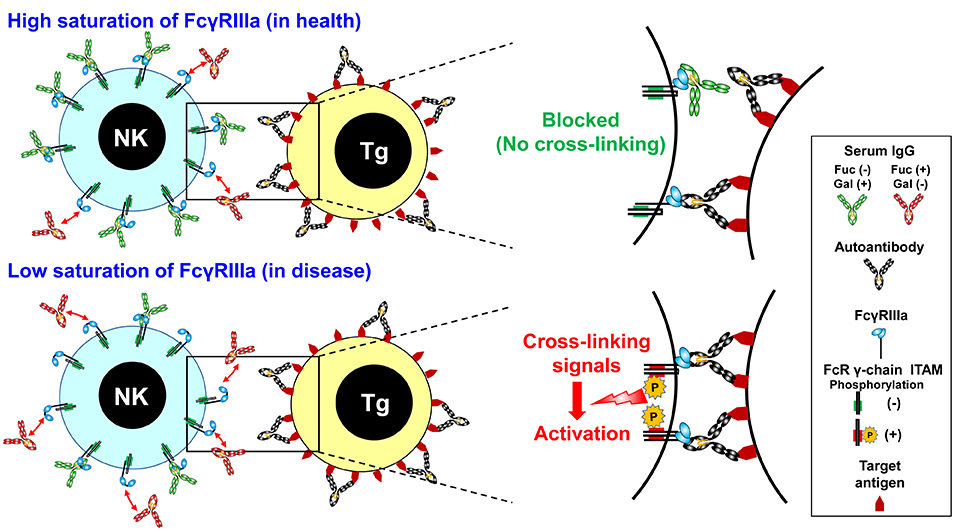
Blockade of activating FcγRs has long been postulated as a possible MOA of IVIG without formal proof. In ITP patients, opsonized platelets are known to be phagocytosed by immune cells that express FcγRIIIa21, and administration of anti-FcγRIIIa antibody results in a rapid increase in platelet counts22,23. In Kawasaki Disease, NK cells of patients respond well to IVIG, resulting in reduction of NK cell proliferation and ADCC activity24. Thus, FcγRIIIa on immune cells is reported to be involved in the pathogenesis of a range of autoimmune/inflammatory diseases25-27 although it remains unknown how IVIG can modulate the functional activities of immune cells. To our knowledge, our study is the first to provide a molecular mechanism underlying the anti-inflammatory effect of IVIG, revealing that galactosylated, nonfucosylated IgG blocks FcγRIIIa cross-linking and inhibit ADCC by NK cells (Fig. 7)4.
Fig. 7. Immunomodulation via FcγRIIIa blockade by galactosylated nonfucosylated IgG in health (Top) and disease (Bottom)
6. Questions on the anti-inflammatory activity of sialylated IgG
The anti-inflammatory properties of sialylated IgG remain controversial. In our animal experiment, homogeneous sialylated IgG glycoforms, regardless of fucose, failed in suppressing inflammation (Fig. 6)4. The working hypothesis on the MOA of sialylated IgG is upregulation of inhibitory FcγRIIb on macrophages in response to TH2 cytokines secreted via the binding of sialylated IgG to the C-type lectin DC-SIGN3,28. However, this theory is not likely valid because DC-SIGN was found not to bind sialylated IgG29,30. Furthermore, because IVIG can exert anti-inflammatory effects in FcγRIIb-deficient mice, it is concluded that FcγRIIb is dispensable for the efficacy of IVIG23,31,32. In fact, the Sambucus Nigra Lectin (SNA)-enriched IgG representing a sialylated IgG in previous studies contains a high proportion of monosialylated IgG species (Fig. 4)3,33. The sialic acid residue of monosialylated IgG is preferentially attached to the α(1-3) arm; therefore, galactose is supposed to be present at the nonreducing end of the α(1-6) arm34. Galactose on the α(1-6) arm makes numerous contacts with amino acid residues of the CH2 domains (Fig. 1), contributing to the stability and Fc effector functions35. Thus, the effect of the SNA-enriched IgG does not necessarily reflect the properties of sialylated IgG alone.
7. Conclusions
The MOA of IVIG has been a long-standing question. We have demonstrated its anti-inflammatory mechanism quantitatively that involves FcγRIIIa blockade by the (G2)2 glycoform, a component of IVIG, thereby inhibiting cytotoxic activity of immune cells. Although further studies are needed to confirm its safety and efficacy, development of the (G2)2 IVIG for therapeutic use may lead to the reduction of the doses of IVIG, thereby decreasing the plasma consumption (Patent pending)36. Additionally, the potent anti-inflammatory (G2)2 IVIG may be a promising treatment option for IVIG non-responders of Kawasaki Disease, etc37.
References
Joao C, Negi VS, Kazatchkine MD, Bayry J, Kaveri SV. Passive Serum Therapy to Immunomodulation by IVIG: A Fascinating Journey of Antibodies. J Immunol 2018;200(6):1957-1963.
Debre M, Bonnet MC, Fridman WH, Carosella E, Philippe N, Reinert P, et al. Infusion of Fc gamma fragments for treatment of children with acute immune thrombocytopenic purpura. Lancet 1993;342(8877):945-949.
Kaneko Y, Nimmerjahn F, Ravetch JV. Anti-inflammatory activity of immunoglobulin G resulting from Fc sialylation. Science 2006;313(5787):670-673.
Mimura Y, Mimura-Kimura Y, Saldova R, Rudd PM, Jefferis R. Enhanced Immunomodulatory Effect of Intravenous Immunoglobulin by Fc Galactosylation and Nonfucosylation. Front Immunol 2022;13:818382.
Mimura Y, Jefferis R. Human IgG glycosylation in inflammation and inflammatory disease. In: Barchi JJ, editor. Comprehensive Glycoscience. Vol. 5. Oxford: Elsevier, 2021:215-232.
Mimura Y, Saldova R, Mimura-Kimura Y, Rudd PM, Jefferis R. Importance and Monitoring of Therapeutic Immunoglobulin G Glycosylation. Exp Suppl 2021;112:481-517.
Mimura Y, Church S, Ghirlando R, Ashton PR, Dong S, Goodall M, et al. The influence of glycosylation on the thermal stability and effector function expression of human IgG1-Fc: properties of a series of truncated glycoforms. Mol Immunol 2000;37(12-13):697-706.
Mimura Y, Sondermann P, Ghirlando R, Lund J, Young SP, Goodall M, et al. Role of oligosaccharide residues of IgG1-Fc in Fc gamma RIIb binding. J Biol Chem 2001;276(49):45539-45547.
Krapp S, Mimura Y, Jefferis R, Huber R, Sondermann P. Structural analysis of human IgG-Fc glycoforms reveals a correlation between glycosylation and structural integrity. J Mol Biol 2003;325(5):979-989.
Shields RL, Lai J, Keck R, O'Connell LY, Hong K, Meng YG, et al. Lack of fucose on human IgG1 N-linked oligosaccharide improves binding to human Fcgamma RIII and antibody-dependent cellular toxicity. J Biol Chem 2002;277(30):26733-26740.
Shinkawa T, Nakamura K, Yamane N, Shoji-Hosaka E, Kanda Y, Sakurada M, et al. The absence of fucose but not the presence of galactose or bisecting N-acetylglucosamine of human IgG1 complex-type oligosaccharides shows the critical role of enhancing antibody-dependent cellular cytotoxicity. J Biol Chem 2003;278(5):3466-3473.
Ferrara C, Grau S, Jager C, Sondermann P, Brunker P, Waldhauer I, et al. Unique carbohydrate-carbohydrate interactions are required for high affinity binding between FcgammaRIII and antibodies lacking core fucose. Proc Natl Acad Sci U S A 2011;108(31):12669-12674.
Mizushima T, Yagi H, Takemoto E, Shibata-Koyama M, Isoda Y, Iida S, et al. Structural basis for improved efficacy of therapeutic antibodies on defucosylation of their Fc glycans. Genes Cells 2011;16(11):1071-1080.
Woof JM, Burton DR. Human antibody-Fc receptor interactions illuminated by crystal structures. Nat Rev Immunol 2004;4(2):89-99.
Kiyoshi M, Caaveiro JM, Kawai T, Tashiro S, Ide T, Asaoka Y, et al. Structural basis for binding of human IgG1 to its high-affinity human receptor FcgammaRI. Nat Commun 2015;6:6866.
van Mirre E, Teeling JL, van der Meer JW, Bleeker WK, Hack CE. Monomeric IgG in intravenous Ig preparations is a functional antagonist of FcgammaRII and FcgammaRIIIb. J Immunol 2004;173(1):332-339.
Patel KR, Nott JD, Barb AW. Primary Human Natural Killer Cells Retain Proinflammatory IgG1 at the Cell Surface and Express CD16a Glycoforms with Donor-dependent Variability. Mol Cell Proteomics 2019;18(11):2178-2190.
Sondermann P, Huber R, Oosthuizen V, Jacob U. The 3.2-A crystal structure of the human IgG1 Fc fragment-Fc gammaRIII complex. Nature 2000;406(6793):267-273.
Wang LX, Tong X, Li C, Giddens JP, Li T. Glycoengineering of Antibodies for Modulating Functions. Annu Rev Biochem 2019;88:433-459.
Tang F, Wang LX, Huang W. Chemoenzymatic synthesis of glycoengineered IgG antibodies and glycosite-specific antibody-drug conjugates. Nat Protoc 2017;12(8):1702-1721.
Ebbo M, Audonnet S, Grados A, Benarous L, Mahevas M, Godeau B, et al. NK cell compartment in the peripheral blood and spleen in adult patients with primary immune thrombocytopenia. Clin Immunol 2017;177:18-28.
Bussel JB. Fc receptor blockade and immune thrombocytopenic purpura. Semin Hematol 2000;37(3):261-266.
Crow AR, Lazarus AH. Mechanistic properties of intravenous immunoglobulin in murine immune thrombocytopenia: support for FcgammaRIIB falls by the wayside. Semin Hematol 2016;53 Suppl 1:S20-22.
McAlpine SM, Roberts SE, Heath JJ, Kasermann F, Issekutz AC, Issekutz TB, et al. High Dose Intravenous IgG Therapy Modulates Multiple NK Cell and T Cell Functions in Patients With Immune Dysregulation. Front Immunol 2021;12:660506.
Finberg RW, Newburger JW, Mikati MA, Heller AH, Burns JC. Effect of high doses of intravenously administered immune globulin on natural killer cell activity in peripheral blood. J Pediatr 1992;120(3):376-380.
Ruiz JE, Kwak JY, Baum L, Gilman-Sachs A, Beaman KD, Kim YB, et al. Intravenous immunoglobulin inhibits natural killer cell activity in vivo in women with recurrent spontaneous abortion. Am J Reprod Immunol 1996;35(4):370-375.
Pradier A, Papaserafeim M, Li N, Rietveld A, Kaestel C, Gruaz L, et al. Small-Molecule Immunosuppressive Drugs and Therapeutic Immunoglobulins Differentially Inhibit NK Cell Effector Functions in vitro. Front Immunol 2019;10:556.
Anthony RM, Kobayashi T, Wermeling F, Ravetch JV. Intravenous gammaglobulin suppresses inflammation through a novel T(H)2 pathway. Nature 2011;475(7354):110-113.
Yu X, Vasiljevic S, Mitchell DA, Crispin M, Scanlan CN. Dissecting the molecular mechanism of IVIg therapy: the interaction between serum IgG and DC-SIGN is independent of antibody glycoform or Fc domain. J Mol Biol 2013;425(8):1253-1258.
Temming AR, Dekkers G, van de Bovenkamp FS, Plomp HR, Bentlage AEH, Szittner Z, et al. Human DC-SIGN and CD23 do not interact with human IgG. Sci Rep 2019;9(1):9995.
Campbell IK, Miescher S, Branch DR, Mott PJ, Lazarus AH, Han D, et al. Therapeutic effect of IVIG on inflammatory arthritis in mice is dependent on the Fc portion and independent of sialylation or basophils. J Immunol 2014;192(11):5031-5038.
Leontyev D, Katsman Y, Ma XZ, Miescher S, Kasermann F, Branch DR. Sialylation-independent mechanism involved in the amelioration of murine immune thrombocytopenia using intravenous gammaglobulin. Transfusion 2012;52(8):1799-1805.
Mimura Y, Katoh T, Saldova R, O'Flaherty R, Izumi T, Mimura-Kimura Y, et al. Glycosylation engineering of therapeutic IgG antibodies: challenges for the safety, functionality and efficacy. Protein Cell 2018;9(1):47-62.
Grey AA, Narasimhan S, Brisson JR, Schachter H, Carver JP. Structure of the glycopeptides of a human gamma 1-immunoglobulin G (Tem) myeloma protein as determined by 360-megahertz nuclear magnetic resonance spectroscopy. Can J Biochem 1982;60(12):1123-1131.
Wada R, Matsui M, Kawasaki N. Influence of N-glycosylation on effector functions and thermal stability of glycoengineered IgG1 monoclonal antibody with homogeneous glycoforms. MAbs 2019;11(2):350-372.
Patent pending (PCT/JP2021/008404) ”Anti-inflammatory non-fucosylated immunoglobulin preparation and method for producing the same”
Shrestha S, Wiener H, Shendre A, Kaslow RA, Wu J, Olson A, et al. Role of activating FcgammaR gene polymorphisms in Kawasaki disease susceptibility and intravenous immunoglobulin response. Circ Cardiovasc Genet 2012;5(3):309-316.