Katsuki Ohtani
Katsuki Ohtani obtained his Ph.D. from Asahikawa Medical University (2003). He started as assistant professor (2004-2012) and has become associate professor (2013-present) in the Department of Microbiology and Immunochemistry, Asahikawa Medical University. His research examines the biological functions of newly discovered collectins.

Nobutaka Wakamiya
Nobutaka Wakamiya graduated from Hirosaki University Medical School (1980) and obtained his Ph.D. from the Graduate School of Medicine, Osaka University (1986). He served as a postdoctoral fellow at the Dana-Farbar Cancer Institute and Harvard Medical School (1986-1987). He was appointed assistant professor in the Department Pathology, Research Institute for Microbial Diseases, Osaka University (1988-2000), after which he moved to Asahikawa Medical University (2000-present) to become professor in the Department of Microbiology and Immunochemistry. His current research focuses on the relationship of collectins with complement-related diseases.
Research on acquired immunity, including cell-mediated and humoral immunity became prominent with advances made in molecular biology in the 1970s. Later the importance of innate immunity became clear with the discovery of the Toll-like receptor. More recently, lectin studies have drawn new attention to immunology. The first study of a carbohydrate chain that led to fusion with biology was the discovery of the mannan-binding protein by Kawasaki and Yamashina. Because this lectin has multimeric structures, it was found to show high binding capacity to the terminal carbohydrate chain of complex carbohydrates and is considered to be a model of pattern recognition of innate immunity. Furthermore, in later years, membrane type lectins such as dectin have been shown to control response to the pathogenic microbe by binding to the carbohydrate chain through the extracellular domain, setting up effective host defense. In this report, we focus on collectin (collagen-lectin) having a collagen-like structure and describe the role of collectin in host defense.
Collectin is a member of the C-type lectin superfamily having a collagen-like structure, and it is classified as five secreted type collectins and one membrane type collectin in Homo sapiens based on the genes and structure of the protein1,2 (Figure 1, Table 1).
(1) MBL (MBP/MBL, mannan-binding protein or mannan-binding lectin)
(2) SP-A (surfactant protein A)
(3) SP-D (surfactant protein D)
(4) CL-L1 (collectin liver 1)
(5) CL-K1 (collectin kidney 1)
(6) CL-P1 (collectin placenta 1)
In the molecular evolution of the collectin genes, 66 genes appear in an amphioxus as the oldest ancestor of the vertebrate2, 3. Fish were found to have four collectin genes, two pulmonary collectin genes appear in land animals, and six collectin genes (COLEC) are present in Homo sapiens (Figure 2). These collectins contain the necessary amino acid sequence in the collagen-like domain called MASP (MBL-associated serine protease)-binding motif to participate in complement activation. This is why this collectin is referred to as a complement-related lectin4 (Figure 3).
The following outlines the main findings dating back to the discovery of collectins. MBL was discovered by Kawasaki and Yamashina in 19805, and in 1989 a report of repeated infection by microbe in MBL-deficient infants by Super et al. revealed the relationship between MBL and innate immunity in human6. MBL presents in the blood, acts as opsonin molecules bound to microorganisms and reduces the number of microorganisms in the body, giving rise to the hypothesis that MBL acts on innate immune function to protect the host. The collectin molecules show opsonic activity and likely play an important role in the body's defense capability. A subsequent genetic study reported that MBL deficiency in humans revealed amino acid alterations and single nucleotide polymorphism (s) (SNPs) in the promoter domain of the MBL gene7 (Figure 4).
Pulmonary collectin SP-A and SP-D were discovered independently after 19858, 9, but the pulmonary collectin gene was not understood at the time because the collectin gene family had not been identified as an MBL. Twenty years after collectins were discovered, three new collectin genes were identified by using reverse genetic technique by Ohtani et al., and the total image of the collectin gene family became clear10-12. The distribution and genome organization of these collectin genes vary greatly. MBL is produced mainly in liver parenchymal cells, and it exists as a secretory protein in blood. Pulmonary collectins SP-A and SP-D are secreted from an alveolus Type II epithelium and a bronchus epithelium, bowel epithelium by an alveolus, a trachea epithelium and an intestinal mucosa and exist primarily as a pulmonary surfactant protein. However, three newly discovered collectins display a unique distribution. CL-P1 was discovered in a placenta cDNA library, but later it became clear that it mainly expressed in the blood vessel tissue of the whole body13. The CL-L1 gene in collectin from liver was cloned and initially reported to be a cytosolic protein, but it later became clear that it was actually a secreted protein, and recently it was revealed that CL-L1 forms a heterodimer with CL-K1 in human blood10, 14. It has been reported that CL-L1 and CL-K1 express in almost all organs of the body, but, because they present in blood, they are considered to be secreted mainly by the liver15. The heteromeric complex of CL-L1and CL-K1 was named CL-LK by Hansen and others, but it is not clear how this complex is formed, and its biological role in human is currently under investigation14.
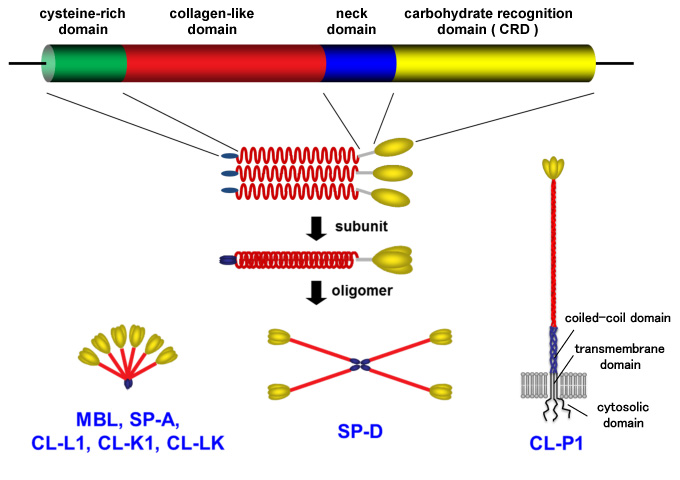
Fig. 1 Molecular structures of collectin
Table 1 Characterization of collectin
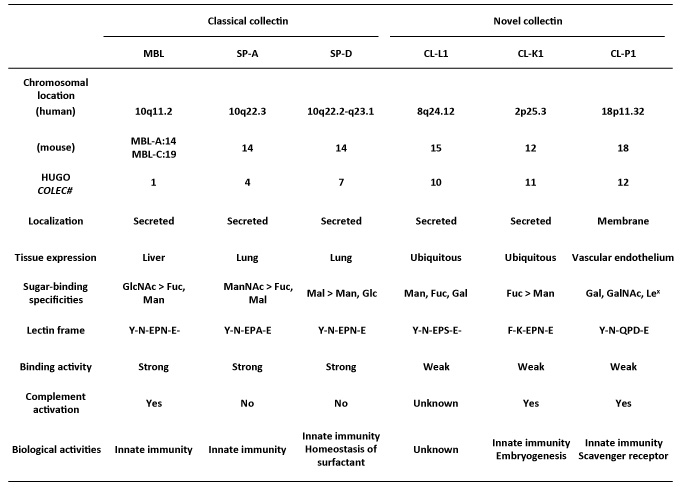
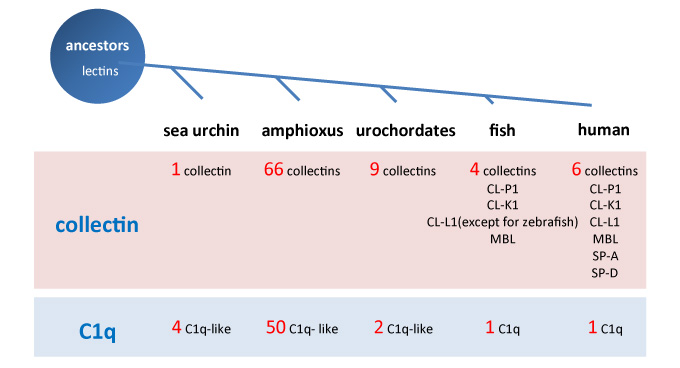
Fig. 2 A schematic of the gene evolution of collectin and C1q

Fig. 3 Alignment of the collagen-like sequence around the putative MASP-binding motif4
XX = any amino acid. ‘:’ entirely conserved. ‘.’ highly conserved. Prolines in the third position in the Gly-X-Y triplet are likely hydroxyprolines. The putative MASP-binding motif in the collagen sequence is indicated in blue. Residues involved in binding are given in bold, and the essential lysine (55 in MBL) is indicated with an asterisk.
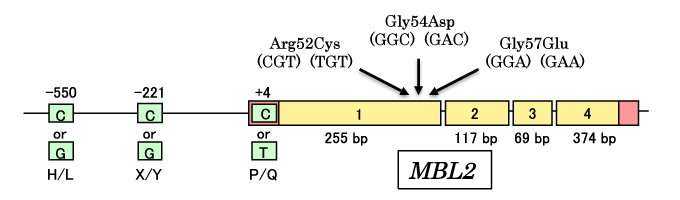
Fig. 4 SNPs of human MBL
The biological defense function of collectins against a variety of microbial infections has become apparent from the molecular biology research and an epidemiological study on MBL deficiency. In Gram-negative bacteria, MBL is believed to recognize the sugar chain structure on the lipopolysaccharide (LPS) of the bacterial surface. The binding with MBL is not seen in Salmonella strains having smooth type LPS, although there is evidence of MBL binding with a Salmonella strain with rough type LPS lacking the O antigen. In addition, MBL binds to the gonococcal LPS, but it is reported that sialic acid added to this carbohydrate chain markedly decreases the binding 17. Furthermore, it has become clear by detailed study using a variant of these bacteria or a carbohydrate chain-related gene knockout strain that the binding site of MBL is the core region of the LPS. On the other hand, the binding site of gram-positive bacteria has been reported to be a lipomannan or glucose-substituted lipoteichoic acid18. MBL shows a higher tendency to bind with gram-negative or –positive bacteria which do not have a capsule than with bacteria having a capsule. MBL binds to these bacteria as an opsonin molecule and is thought to reduce the number of bacteria in cooperation with subsequent phagocytic cells.
Mycobacterium tuberculosis differs from these bacteria in that many sugar structures, including lipoarabinomannan (LAM) and lipoarabinogalactan (LAG) and phenolic glycolipid (PGL), a complex of a long chain fatty acid and a sugar called the cord factor (Trehalose-dimycolate, TDM), exist on the bacteria surface. Among these MBL binds strongly to LAM. The binding of MBL to mannose cap LAM (Man LAM) is higher than to LAM (AraLAM), and it is reported that Man LAM is seen in strongly pathogenic bacteria19. In addition, it has been observed that binding of MBL to bacteria activates complement on the bacterial surface20, and MBL enables complement activation for a glycosyltransferase gene defective Brucella21. It has also been reported that collectin acts to promote rather than defend against acid-fast bacterial infection. In a group of patients suffering from tuberculous meningitis, glycine of the 54th amino acid did not change to glutamine of MBL in comparison with a group not affected22. The 54th amino acid alteration of MBL proved to greatly lower blood concentration of MBL, and it was suggested that the group having a low concentration of MBL failed to contract tuberculous meningitis. These results indicate that collectin has a dual function, infection defense and promotion, in bacterial infections.
Viruses multiply by invading cytoplasm of the host without being able to proliferate independently to reproduce a viral gene of itself. The relationship between collectin and viral infection has been reported for various viruses, but here we take the influenza A virus as an example. In the early 1970s, three kinds of endogenous inhibitors, α, β,γin blood, were found to show hemagglutination inhibition (HI) activity for the influenza virus. Anders and Wakamiya found that the main factor of the β inhibitor was collectin23, 24. Kase et al. subsequently demonstrated that MBL expressed infection inhibitory activity by binding to the sugar chain of two envelope proteins of the influenza virus, hemagglutinin (HA) and neuraminidase (NA)25. Furthermore, MBL binds to the influenza virus acting as an opsonin molecule, not by direct neutralization action, and removes the virus by phagocytosis using neutrophils and macrophages26 (Figure 5).
It is reported that pulmonary collectin SP-D provides anti-influenza properties by a mechanism the same as that of MBL27. In addition, MBL and SP-D have neutralization activity for influenza A virus subtype H1N1 and H3N2, because the two subtype viruses have high mannose type, hybrid type carbohydrate chains in HA and NA. This is consistent with the fact that the binding of collectin with H1N1 and H3N2 viruses is high, and the binding of collectin to subtype H2N2 is low because it has few of these carbohydrate chains25, 28. Furthermore, Kase and Kawai demonstrated that MBL exhibits neutralization activity by the budding inhibition of the virus particle by binding to NA25, 29. In contrast, SP-D does not have the binding capacity to NA and does not show neutralization activity by the budding inhibition. Moreover, in an epidemiological survey by Kase and others, the infection neutralization activity level of MBL was 0.31±0.67μg/ml in 67 subtype H3N2 influenza viruses isolated during the years 1990-1995, but the neutralization level was lower than the human MBL serum level 1-2μg/ml, indicating that freshly isolated strains are sensitive to MBL. However, HI active concentration of more than 2.5μg/ml was also observed around 2% fresh virus isolates, showing the potential that collectin-resistant influenza viruses are present in a seasonal flu virus25. In 2009, there was an outbreak of influenza virus (A/H1N1/pdm) of subtype H1N1, and WHO reported more than 16,000 deaths worldwide. The epidemic occurred because this A/H1N1/pdm virus had not been prevalent for the past 50 years, and most individuals did not carry antibodies against it. When acquired immunity is not effective against an influenza virus, it is thought that natural immunity usually works to protect against infection, but it was reported that this virus showed no sensitivity to collectin such as MBL or SP-D28. Prevalent seasonal influenza virus H1N1 has 3-5 high mannose type and hybrid type carbohydrate chains in HA, but A/H1N1/pdm virus only has one carbohydrate chain of HA and collectin cannot bind to HA, so cannot function as the neutralizing antibody28. In other words, for the influenza virus A/H1N1/pdm, which appeared in 2009, the world population had neither acquired immunity (immunological memory due to the antibody) nor natural immunity (collectin), enabling A/H1N1/pdm outbreaks throughout the world.
There are fewer reports on infection control by collectin for the eukaryotic pathogens such as mold, protozoa, and parasitic worms than there are for bacteria and viruses. There is a report on malaria with observation of the 54th or/and 57th amino acid substitution by the SNP of MBL gene and a strong correlation shown between blood concentration of MBL and severity of malaria30. Further reports indicate the possibility that collectin is important in the defense against infection due to Aspergillus-caused pneumonia31 and Cryptosporidium-caused diarrhea32.
Against serious microbial infectious diseases that threaten human life, the first priority of the host is to reduce the number of microbes and elimination of the microbe by opsonic activity using collectin is the most primitive and initial action. When opsonic activity of collectin is not sufficient for host defense, a mechanism activating the complement system is available to protect life. It is known that collectin has the ability to activate complement using the lectin pathway as well as its neutralization and opsonization capabilities against microbes (Figure 6). This complement activation can sometimes be very strong, thus it can serve not only to eliminate the microbe but also cause excessive local or systemic inflammation and seriously damage host tissue. In mouse infection experiments using influenza virus by Ling et al., A/H1N1/pdm and avian H9N2 showed MBL binding activity but there was no effect on lung damage caused by the virus infection33. MBL wild type mouse is shown to be stronger against lung tissue damage than MBL KO (knock out) mouse. Cytokine production by influenza virus infection is remarkably higher in this MBL wild type mouse than in KO mouse, and increased cytokines may cause deterioration in general physical condition, weight loss and serious tissue injury.
The function of MBL during organ transplantation has been described in terms of complement activation. More than 2000 lung transplantations are performed every year throughout the world, and it has become a common treatment for end-stage lung disease. The first year after transplantation is the most important for survival, and advances in surgical procedures and progress in immunosuppressive agents have significantly increased the survival rate. Pulmonary dysfunction may occur early due to ischemia-reperfusion, acute rejection and bacterial infection after transplant. Three months after transplantation infectious diseases due to fungus, virus, and mycobacteria, occlusive bronchiolitis syndrome (BOS) and chronic rejection become serious problems. Munster et al. discovered a correlation between BOS onset after transplantation and the promoter region that defines the concentration of MBL, and observed that SNPs of the collagen-like region were related to lung transplant survival34. Regarding BOS, only the recipient of the lung of a donor having an X-allele showed good prognosis of graft survival after transplant. In this case, a precise co-relation between blood MBL concentration and MBL genotypes was not found, and the transplanted donor lung fares better with less MBL produced. These factors appear to determine the transplant prognosis and suggest the possibility of suppressing injury and inflammation of lung tissue34. These findings indicate that in transplanted lung tissue, complement activation function of MBL might not be beneficial.
Similarly, in renal transplantation, data indicating that a lower concentration of MBL in the donor is useful for successful transplant have been reported in an epidemiological survey on mouse and human35. However, in liver and heart transplantation, patients with low MBL can easily progress to acute rejection by serious infection and post-transplant coronary artery disease36, 37. Generally, collectin works to defend against pathogenic infection by natural immunity and lead the living body to a normal state, but when excessive complement activation occurs, the reaction causes serious damage to the host and is considered to be a risk factor.
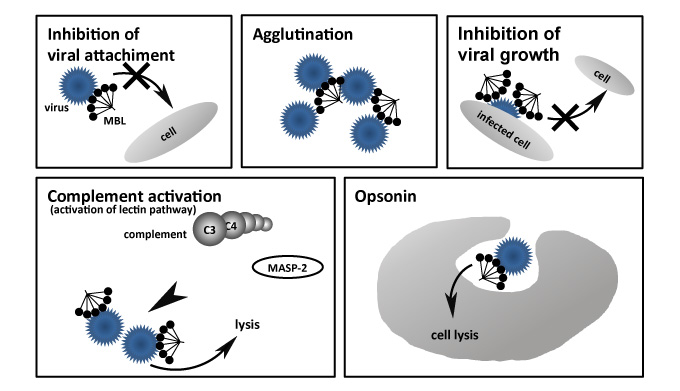
Fig. 5 Anti-viral functions of collectin

Fig. 6 Complement activation pathway
CL-P1 is the only membrane protein in the collectin group. Fukuda and colleagues cloned the zebrafish CL-P1 gene and examined function of CL-P1 at the individual level38. Zebrafish CL-P1, as well as in mouse and human tissue, is primarily seen in vascular tissue. Loss of the dorsal aorta and intersegmental artery, as well as a remarkable abnormality of the trunk formation with pericardium edema were observed, and the possibility that CL-P1 expression participates in the morphosis of the fish embryo was clarified with the CL-P1 gene knockdown zebrafish embryo using morpholino oligonucleotide38. Furthermore, in CL-P1 gene knockdown embryo, decrease of vascular endothelial growth factor (VEGF) mRNA was observed, and CL-P1 may play an important role in blood vessel construction in early development because recovery of the abnormal morphology by co-administration of VEGF mRNA was observed, suggesting the existence of a VEGF and CL-P1 related pathway.
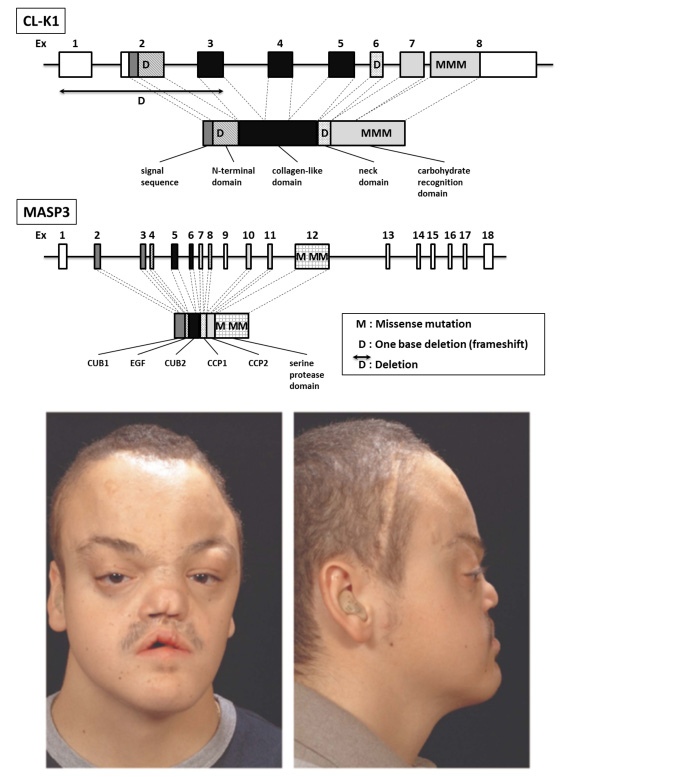
Rooryck and others reported that gene abnormality of either CL-K1 or MASP-3 (MBL- associated serine protease-3) is involved in the autosome recessive hereditary disease called the 3MC (Carnevale, Mingarelli, Malpuech, and Michels) syndrome39 (Figure 7). 3MC syndrome is characterized by various abnormalities, e.g., hypertelorism, blepharophimosis, blepharoptosis, craniosynostosis, cleft palate, mental retardation, sexual organs hypoplasia. These occur in most patients as a result of consanguineous marriage, and the frequency of this hereditary disease is extremely low, but there exists a possibility that complement activation itself by collectin CL-K1 and MASP3 is important to the morphogenesis of the whole body of humans. In this disease, loss of blood CL-K1 due to the CL-K1 gene abnormality has been confirmed. It is thought that there is no CL-LK in 3MC patients because most CL-K1 molecules exist in blood as CL-LK. However, there is no report of gene abnormality of CL-L1 in this disease, and at present it is unknown whether independent CL-K1 is connected with MASP-3 protein, or CL-LK participates at the time of fetal morphogenesis. In addition, Takahashi et al. found increased levels of CL-K1 in blood in American disseminated intravascular coagulation (DIC) patients40. DIC is a multiple organ dysfunction syndrome and a critical disease with a high death rate. Early treatment is important, but no useful biomarker currently exists. This is the first report that showed a relationship between blood CL-K1 level and DIC, but the mechanism is unknown and further study is necessary. Three newly identified collectins, CL-P1, CL-K1, and CL-L1, are different from those that protect against infection and the body's defense mechanism against conventional microorganisms. They may play an important role in complex systems such as the complement system and the coagulation and fibrinolysis system. It is also assumed to be important in tissue homeostasis.
Among collectins, SP-A and SP-D, which are pulmonary collectins, are exceptional collectins that do not activate a complement system. Direct opsonin and neutralization effect is their main function and it is possible that a limited function exists that may cause them to lose their complement activation ability. Pulmonary damage occurs to the entire lung, leading to respiratory failure when the lungs show excessive immunoreaction because it is always exposed to the outside world. However, the complement-related lectins, except SP-A and SP-D, appear to be set in complement activation always acting integrally at a local site. CL-P1 is not the exception in complement activation, but secretion type CL-P1 attaches directly to the microbe surface and induces complement activation 41. Roy et al. very recently demonstrated a new complement activation pathway in which CL-P1 traps CRP, acute inflammation protein, at the cell surface and starts a classic pathway, activating an amplification loop as an alternate pathway42. CL-P1, which functions as a scavenger receptor, has been described above. It is clear that CL-P1 is involved in the complement system and it is possible that the complement system is connected with collectin in a more complicated manner than is currently known. Research on collectin as a complement-related lectin has begun to elucidate its role in the living body (Figure 8).
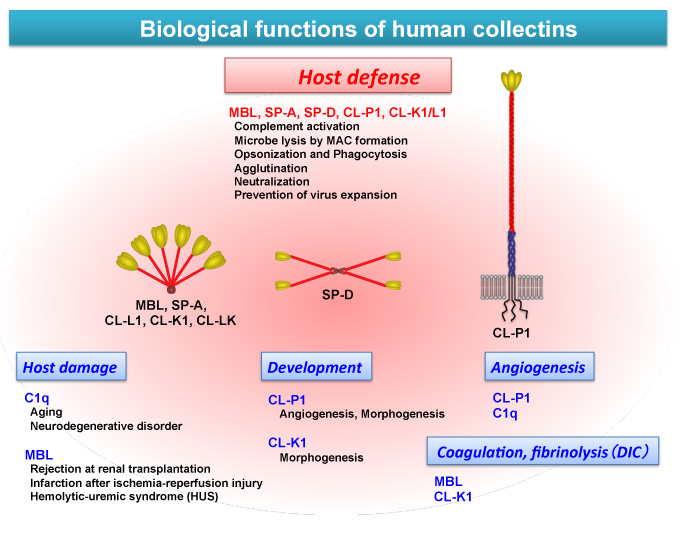
Fig. 8 Biological functions of human collectins
Acknowledgment
We thank all our collaborators at the Research Institute for Microbial Diseases, Osaka University, Osaka Prefectural Research Institute, and Asahikawa Medical University. Wakamiya sincerely thanks the late Professors Shiro Kato, Masaharu Naiki, and Ikuo Yamashina for their special support.