
Yasuo Suzuki
Professor Suzuki graduated from the Shizuoka College of Pharmacy, Graduate School of Pharmacy, acquiring his Ph.D. in 1974. He was appointed as the dean and post graduated dean of the University of Shizuoka, School of Pharmaceutical Sciences on 1996-2002. He served as the vice president of Japanese Society of Carbohydrate Research. He was awarded The Pharmaceutical Society of Japan award 2004 and Chunichi Cultural Award 2004. He occupies a new post as professor in the Life Health Department, specializing in biochemistry, virology and glycobiology with focus on the elucidation of the carbohydrate chain function in viral infections and development of relevant drugs. He is also the Adjunct professor of Griffith University, Australia, and visiting professor of Zhejiang Academy of Medical Science, China. He organized the sialoglycosceince 2006, 5th International conference, Mishima, Japan(Aug.27-30,2006).
Research on influenza virus (Fig. 1) receptors has been ongoing for many years. In the mid twentieth century, the receptor destroying enzyme neuraminidase was found in influenza virus particles, and sialic acid (Sia)-containing sugar chains were recognized as being receptors of influenza virus 1-3. The precise chemical structures of the receptor sialosugar chains were determined by Suzuki et al. 4-6.
The natural host of the influenza A virus which caused a world-wide pandemic is wild water fowl. Influenza virus can be transmitted to a domestic intermediate host, e.g., chickens and pigs by contact with infected wild water fowl that migrate worldwide. The host range mutation of influenza virus from bird to human and change in host receptor binding specificity of the virus are closely related. In 1997, there was an outbreak of a highly pathogenic avian influenza virus (H5N1) in Hong Kong. This virus is lethal to chicken and continuously spreading with a high lethality rate (approx. 60%) to humans. In 2009, a new influenza virus (H1N1) containing virus genes of avian, human, and swine influenza caused a pandemic from Mexico. In February 2013, a new type of avian influenza virus (H7N9) was found in Shanghai. This virus is weakly pathogenic to chicken. However, to date, infection is spreading to humans with high pathogenicity and lethality rate of over 25%. How does the influenza virus enter the human body and acquire the ability to spread? The change in host receptor binding specificity of the virus is closely related to the elucidation of this mechanism.
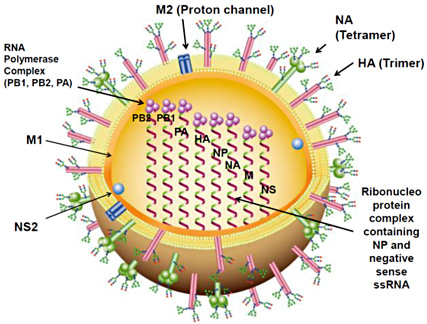
Fig. 1 Schematic diagram of an influenza A virus
Two surface glycoproteins, hemagglutinin (HA) and neuraminidase (NA), and the M2 proton channel are present in the viral envelope, which is derived from the host cell membranes. Glycans on the HA and NA are also synthesized from the host cell glycosylation system. The ribonucleoprotein complex comprises a viral RNA segment associated with the nucleoprotein (NP) and three RNA polymerase proteins (PA, PB1, PB2). The matrix (M1) protein is associated with both nucleoprotein and the viral envelope. Non-structural protein 2 (NS2) is also present in the virion. The eight viral negative strand RNA segments produce 12 proteins, PB1, PB1-F2, N40, PB2, PA, HA, NP, NA, M1, M2, NS1, NS2.
Influenza is one of the most widely distributed zoonosis in the world. The host range of the influenza virus is shown in Fig. 2. Sixteen subtypes of influenza A virus hemagglutinin (HA) spike have been identified. The first step in influenza infection of human respiratory tract is binding of the virus hemagglutinin (HA) glycoprotein to sialic acid-terminated receptors. The binding of different strains of virus for the receptor is determined by α-linkage of Sia to Gal (galactose). Sixteen subtypes (H1-H16) of the HA of influenza A viruses are currently known. Sialyl sugar chains on the host cell membranes are expressed as glycoproteins (N- and O-glycans), sphingoglycolipids (gangliosides), GPI anchors and mucins 7-9. More than 50 molecular species of sialic acids (Sias) are known in nature; among them, N-acetylneuraminic acid (Neu5Ac) and N-glycolylneuraminic acid (Neu5Gc) are important for the infection of influenza A, B viruses. HAs of avian influenza virus strains preferentially bind to oligosaccharides that terminate with Sia linked to galactose by α2-3-linkages (Siaα2-3Gal; avian-type receptor), whereas the HAs of human influenza virus strains prefer oligosaccharides that terminate with a Sia linked to Gal by α2-6-linkages (Siaα2-6Gal; human-type receptor) 8,9.
The shift from Siaα2-3Gal binding to Sia α2-6Gal-binding specificity is a critical step in the initial adaptation of avian influenza viruses to human hosts. Actually, all the pandemic strains of the 1918 Spanish influenza (H1N1), 1957 Asian flu (H2N2), 1968 Hong Kong flu (H3N2), as well as the recent pandemic H1N1 2009 virus, exhibited human-type receptor-binding specificity 10-13 although their HAs originated in non-human species.
Sialyl sugar chain-specific lectin staining of tissue sections has revealed that epithelial cells in the human upper respiratory tract predominantly express Siaα2-6Gal, whereas lower human respiratory tract and lungs express both human-type (Siaα2-6Gal) and avian-type (Siaα2-3Gal) receptors 14. Human and avian-type receptors have also been detected in pig respiratory tract 15. In aquatic birds, including ducks and geese, avian-type receptors dominate in tracheal epithelial cells 16-18. However, in terrestrial birds, including chicken, turkey, and quail, both avian-type (Siaα2-3Gal) and human-type (Siaα2-6Gal) receptors are detected in the epithelial cells 16, 19, suggesting that these species can support the replication of both avian and human influenza viruses and act as adaptation hosts for receptor switching of avian strains. Notably, some H5N1 and H9N2 viruses isolated from terrestrial birds, such as chicken and quail, bind to human-type receptors 20, 21.
The receptor binding specificity of the influenza viruses described above was determined by several methods, including agglutination assays using sialosugar-modified-erythrocytes, solid-phase binding assays using native and synthetic sialosugar chains, and recent glycan microarray assays 8,9,12,22. Mass spectrometry and sialic acid-specific lectin staining of tissue sections have revealed the structure of N-sialoglycans of trachea and lung of pig 23,24, digestive tract of quail and chicken 25. Recently, N- and O-glycan composition of the human lung, bronchus and nasopharynx has been characterized by mass spectrometry 26.
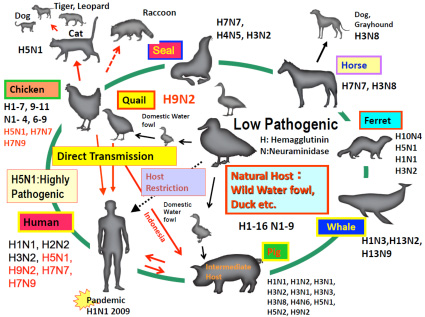
Fig. 2 Host range of influenza A viruses
The native host of influenza A viruses (subtypes: H1-H16, N1-N9) is wild waterfowl such as duck, which is the origin of all the influenza A viruses on earth. The virus originally had low pathogenicity but acquired high pathogenicity over time by transmission to and propagation among domestic fowl chicken on land. Influenza viruses mutate easily, acquiring high pathogenicity and expanding their host range. H5N1, H7N7, H7N9 viruses have been transmitted from chicken to humans and H9N2 virus from quail to humans, but these viruses are not transmitted efficiently among humans.
The present author and his research team have discovered isolates of highly pathogenic avian influenza (H5N1) that have acquired the ability to bind to human-type receptors in infected humans. In other words, not only have we discovered avian-type receptors in humans infected by H5N1, but also isolates that have acquired the ability to bind to human-type receptors in Hong Kong 27, Thailand 28, and Vietnam 29. We 30 recently found a strain isolated from a highly pathogenic avian influenza H5N1 strain taken from a chicken in Egypt that can bind to human-type as well as avian-type receptors. Nidom et al. 31 found that one of the isolates of H5N1, which is propagating at a significant rate in Indonesian pigs (13% of surveyed pigs), had both avian-type and human-type receptor binding. A strain that can bind to avian-type and human-type receptors has also been reported in highly pathogenic H7 isolates of chicken and turkey 32, as well as H9 strain isolated from quail 21. This result shows that highly pathogenic avian influenza (H5N1) and other subtype avian influenza viruses acquire human-type receptor binding specificity during circulation among livestock such as chicken, quail, turkey, and pig.
Human-type receptors were found in the upper respiratory tract, and both human-type and avian-type receptors were detected in the lower respiratory tract and alveoli of humans14. Therefore, if an avian virus reaches the lower respiratory tract and alveoli of a human, an infection mediated by an avian-type receptor may cause serious pneumonia. In this case, HA mutations that randomly occur in the viral replication process within the human respiratory tract may generate a new mutant adaptive to human within the human body when a strain that has acquired adaptability to human-type receptor (Neu5Acα2-6Gal) existing mainly in the human respiratory tract is selected. Structural analyses of N-sialoglycans of pig respiratory tract and the digestive tract of quails and chickens revealed the presence of human-type as well as avian-type sialosugar chains in the virus target organs of these animals 23,25. These results strongly suggest that human-type receptor binding specificity can be generated not only in human but in terrestrial livestock such as chicken and quail. The H5N1 strain which can bind to both avian and human-type receptors are thought to raise the possibility of human-to-human transmission because they can bind to human-type receptors that exist in the throat and upper respiratory tract of human.
In the case of the influenza virus (H1N1) found in Spain in 1918, it was discovered that only two amino acid substitutions within a hemagglutinin molecule caused the avian-type receptor binding to change to human-type receptor binding. In other words, the first wave viruses HA, E190D, 225G were avian and human-type receptor binding, whereas the second wave viruses HA, 190D, G225D bound to human-type receptors only. This mutation is thought to have made human-to-human upper respiratory tract infection easier. Another subtype, 2009 pandemic H1N1, also spread quickly throughout the human world, but this virus HA also had 190D, 225D mutations, which had acquired the property to bind only to human-type receptors 33-35.
In the case of the pandemic Asian flu virus (H2N2 subtype) of 1957, there were three receptor binding specificities, avian-type (226Q, 228G), human-type (226L, 228S), and an intermediate type (226L, 228S) of human isolate HAs in the initial stage. However, gradually all H2 viruses acquired binding specificity (226L, 228S) to human-type receptors only 36.
The avian influenza (H7N9) outbreak in February 2013 infected 132 people by May 2013, 37 of whom died. This virus showed a more rapid infection to humans compared to H5N1, but human-to-human transmission has not been confirmed to date. The receptor binding specificity of this virus has been published 37-39. All viruses isolated from the target humans indicated binding to both avian and human-type receptors, and also showed the acquisition of a binding specificity to extended N-glycans 40, which are reported to be present in human respiratory tract. The human seasonal virus (H1N1) that was used as control and the 2009 pandemic H1N1 virus preferentially bind to human-type receptors, whereas duck virus and highly pathogenic avian influenza H5N1 preferentially bind to avian-type receptors. This virus is weakly pathogenic to chicken but is highly pathogenic to human. H7N9 virus can bind to both avian-type and human-type receptors, and can invade epithelial cells in the human lower respiratory tract and type II pneumonocytes in alveoli. This virus also replicated efficiently in ex vivo lung and trachea explant culture and several mammalian cell lines. These results indicate that the H7N9 virus poses a potentially high risk to humans 38.
Kawaoka, Fouchier and their colleagues 41-44 revealed that experimental substitution of a few amino acids in avian H5 hemagglutinin confers acquisition of human-type receptor binding specificity and respiratory droplet transmission in ferrets, which are used as animal models of human influenza infection. Imai et al., 41 identified a reassortant H5 HA/H1N1 virus --comprising H5 HA (from an H5N1 virus) with four mutations (N224K, Q226L, N158D, T318I) and the remaining seven gene segments from a 2009 pandemic H1N1 virus-- that was capable of droplet transmission in a ferret model. Herfst et al., 43 genetically modified native A/H5N1 virus by site-directed mutagenesis and subsequent serial passage in ferrets. The mutated H5N1 virus gained the ability to bind to human-type receptor and to be transmitted by aerosol or respiratory droplets (airborne transmission) between ferrets. This virus had four amino acid mutations (Q226L, G228S, T160A, H107Y). Both studies demonstrated that amino acid substitution in 220-loop (N224K, Q226L in 41, and N224K, G228S in 43) and lack of 158-161 glycosylation near the receptor binding pocket in highly pathogenic avian H5 hemagglutinin as well as amino acid substitution (T318I in 41; H107Y in 43) in the stalk region of the H5 HA, spike are critical to shift the receptor binding specificity and droplet transmission between ferrets. T318I or H107Y mutation in stalk region, which is distant from the receptor binding pocket of the H5 HA may be critical for the structural stability of H5 molecule 41, 43.
Lack of the highly pathogenic avian H5N1 hemagglutinin HA 158-161 (H3 numbering) glycosylation site appears to be critical for H5 virus transmission in mammals. Avian H5N1 viruses lacking the HA 158-161 glycosylation site appear to transmit to human more readily than those that possess the glycosylation site, at least in the genetic background of Egyptian viruses. Both studies and findings by others 45-47 suggest that a shift towards human-type receptor-binding specificity from avian-type receptor specificity may be necessary, but not sufficient, for H5N1 virus transmissibility in mammals.
All known pandemic influenza viruses originating in avian strains finally acquired preferential binding specificity to human-type receptors (Neu5Acα2-6Gal). Intermediate strains that bind to both avian- and human-type receptors sometimes appear in the early stage of outbreak. Current avian H5N1 and H7N9 influenza A viruses occasionally infect humans and some of them acquire binding specificity to human-type receptors. H7N9 strains in particular were found to infect humans, causing rapidly progressing severe respiratory tract infection and mortality. But neither H5N1 nor H7N9 virus is currently transmitted efficiently among humans.
Through a database search, Newmann et al. 48 identified two H5N1 viruses, A/Muscovy/duck/Vietnam/NCVD-11/2007 and A/duck/Egypt/10185SS/2010, that encode HA-220K and have lost the HA154-156 glycosylation site, indicating that only two additional mutations are needed to create variants with the transmissibility features identified in the Kawaoka group study.
In the future, further monitoring and research including 1) monitoring of the binding specificity not only to Neu5Acα2-6Gal, the two non-reducing terminal sugar chains of the sialoglycan receptors, but also to a more complex and extended human-type receptors (extended sialyllactosamine sugar chains, Neu5Acα2-6 (Galβ1-4GlcNAc)nβ1- : n=1-3) that may exist in the human upper respiratory tract 49 and 2) monitoring of not only the HA mutation, but also the activity balance with the neuraminidase related to viral propagation and budding from the host cells 50 are necessary. Finally, the development of a highly precise, sensitive, simple and high through-put field monitoring system that can survey the adaptation of the receptor specificity of avian H5N1 and H7N9 viruses to the human-type receptors present in animals and humans is important.



