
Takashi Angata
Dr. Takashi Angata graduated from the Faculty of Science, University of Tokyo, in 1993 and obtained his Ph.D. degree in biochemistry from the Graduate School of Science, University of Tokyo, in 1998.
He has served as Post-doctoral Fellow to Professor Ajit Varki at the University of California San Diego (1998-2003), Research Scientist at the National Institute of Advanced Industrial Science and Technology in Japan (2003-2009), Specially Appointed Associate Professor at Osaka University (2009-2011), and now as Team Leader at RIKEN Advanced Science Institute (2011-present). The author has contributed to the expansion of the Siglec family of sialic acid recognition proteins, and made several seminal discoveries in the field, including the concerted evolution of activating and inhibitory Siglecs as paired receptors. His current research is focused on the role of activating Siglecs in innate immunity and immunity-associated diseases.
All living cells are coated with glycoconjugates, with the glycans constituting the outermost layer of each cell. Because of their location, glycans are highly suitable for interacting with various agents from both inside and outside the body of multicellular organisms. Pathogens of various classes, i.e., viral, bacterial, and eukaryotic pathogens, have developed many ways to utilize glycans, as described in other articles of this series.
This article introduces a fledgling field of study on the interactions between pathogens and Siglecs, a family of sialic acid (Sia)-recognition proteins in vertebrates. A brief overview of Siglecs and their endogenous functions including examples of Siglec-pathogen interactions (Table 1) is followed by a discussion on the possible origin of activating-type Siglecs. This article concludes with perspectives on future directions of study.
Siglecs are a family of Sia-recognition proteins belonging to the immunoglobulin superfamily 1,2. Most Siglecs are expressed on the cells of the immune system and involved in the regulatory signaling of the cells that express them (Fig. 1). Based on the facts that (i) most Siglecs are expressed on the cells in the innate immune system, i.e., the cells that respond to "non-self" in the early phase of immune reaction, (ii) most Siglecs have inhibitory signaling properties, and (iii) the expression of Sias (i.e., Siglec ligands) is mostly restricted to the deuterostomes (the branch of animals that includes vertebrates, echinoderms, etc.), it has been postulated that the primary function of Siglecs in the immune system is to recognize Sias as a signature of "self " ( "self-associated molecular pattern" or SAMP, in opposition to "pathogen-associated molecular pattern" or PAMP) and to dampen the autoimmune reactions that could develop against the body痴 own cells 3. If this hypothesis is true, then it can be predicted that some pathogens will exploit this property of Siglecs to subvert host immune responses against them. There are in fact some pathogens that appear to do this, as described below.
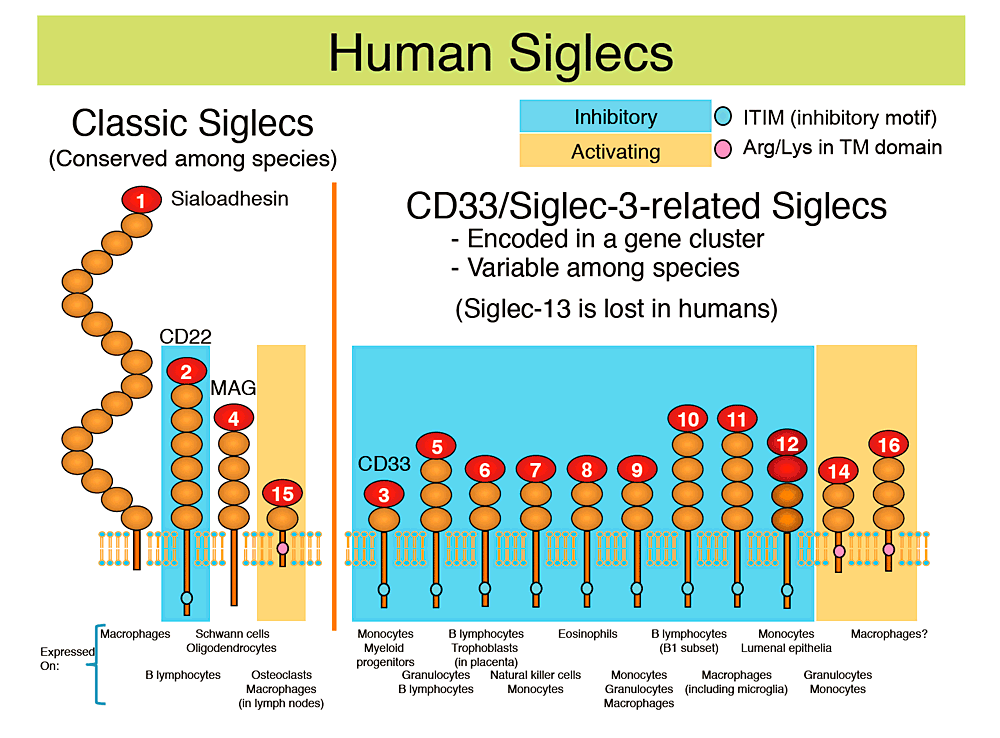
Fig. 1 Human Siglecs
Human Siglecs are shown with cartoon diagrams (each oval represents an immunoglobulin-like domain), and the representative cell types that naturally express them are indicated below. Inhibitory-type Siglecs (which have an immunoreceptor tyrosine-based inhibitory motif, or ITIM, and recruit tyrosine phosphatase SHP-1) are shaded with pale blue background, and activating-type Siglecs (which have a positively charged amino acid residue in the transmembrane domain and interact with an adapter protein called DAP12, which in turn recruits tyrosine kinase Syk) are shaded with orange background.
Human Siglecs can be also classified into two groups based on their patterns of evolution; one group is "classic Siglecs", which are conserved among species; the other group is "CD33-related Siglecs", which are encoded in a gene cluster and show a higher degree of sequence similarity to each other than to the members of "classic Siglecs". The latter group shows lower degree of conservation among species.
Biosynthesis of Sias requires an elaborate series of reactions and is metabolically costly. Despite this fact, a small number of bacteria, mostly those associated with human diseases, express Sias 4. The expression of Sias by these bacteria, then, should confer some advantage on them over those do not express Sias. Complement factor H, which binds to the surfaces covered with Sias and/or heparan sulfate and negatively regulates the Alternative Pathway of the complement system to protect "self" cells/tissues from complement-mediated attacks 5, is considered to be the selective force that provides an advantage for those bacteria that express Sias. Sias may also mask underlying glycans that could elicit antibody responses. In addition to these mechanisms, it appears that "interaction with Siglecs" is also a part of the picture.
Some alternative ways by which bacteria engage Siglecs have been identified so far (Table 1 and Fig. 2): (I) synthesize Sias on their own to interact with Siglecs; (II) borrow Sias from the host to interact with Siglecs; and (III) express protein that directly interacts with Siglecs.
The first category of bacteria (that includes Neisseria meningitidis, Campylobacter jejuni, and group B Streptococcus type III) employs the strategy that is most costly, i.e., synthesize Sias on their own, while the outcomes appear to be rather diverse. In the case of N. meningitidis (a gram-negative bacteria that causes meningitis), interaction with sialoadhesin/Siglec-1 and Siglec-5 enhances phagocytosis by macrophages and may be detrimental to them 6. C. jejuni (a gram-negative bacteria that causes food poisoning, and in rare cases a neurological disorder called Guillain-Barré syndrome) interacts with Siglec-7 7 or sialoadhesin/Siglec-1 8, depending on the terminal structure of their lipooligosaccharides (LOSs). LOS of C. jejuni either have terminal Neu5Acα2-3 or Neu5Acα2-8 residues, and the preferential binding of Neu5Acα2-8+ bacteria with Siglec-7, which is expressed on immature dendritic cells, influences the expression of dendritic cell proteins involved in the polarization of helper T cells 9. In the case of group B Streptococcus (GBS; a gram-positive bacteria that occasionally causes meningitis or septicemia in newborns) type III, it has been reported that the Sia-dependent interaction between the bacteria and Siglec-9 markedly attenuates the responses of neutrophils against the bacteria 10.
The second category of bacteria that includes Pseudomonas aeruginosa adsorbs sialylated glycoproteins from the bodily fluid of the host. P. aeruginosa (a gram-negative bacteria that causes various diseases in immnocompromised hosts) that has adsorbed host plasma glycoproteins engages Siglec-9 and attenuates inflammatory responses of neutrophils 11. My colleagues and I recently found that Haemophilus influenzae (a gram-negative bacteria that occasionally cause otitis media and pneumonia), which scavenges free Sias from human body and utilize them to decorate its LOSs 12, can interact with some Siglecs (unpublished observations). It may be another example of bacteria that borrow Sias from the host to interact with Siglecs, although the significance of such interactions in the context of immune regulation is not yet known.
The third category of bacteria that includes GBS type Ia expresses a protein that directly engages Siglecs by protein-protein interaction. GBS type Ia express β protein that engages Siglec-5 in a Sia-independent manner, and this interaction attenuates innate immune responses by granulocytes 13.
Of course these are just a few examples, and most of the possible Siglec interactions with bacteria and their consequences have yet to be studied.
Table 1 Pathogens reported to interact with Siglecs
| Pathogen | Siglec involved*1 | Pathogen molecule involved | Outcome | Ref. | |
| Virus | |||||
| Porcine reproductive and respiratory syndrome virus (PRRSV) | Sialoadhesin/Siglec-1 (pig) | Sialic acids on GP5 envelope glycoprotein | Enhanced infection | (15, 29) |
|
| Human immunodeficiency virus (HIV) | Sialoadhesin/Siglec-1 | Sialic acids on gp120 envelope glycoprotein | Enhanced infection | (16, 17) |
|
| Varicella zoster virus (VZV), herpes simplex virus (HSV) | Myelin-associated glycoprotein/Siglec-4 | Glycoprotein B (involvement of sialic acids is not reported) | Enhanced infection | (20) |
|
| Bacteria | |||||
| Neisseria meningitidis | Sialoadhesin/Siglec-1 Siglec-5 | Sialic acids on LPS | Enhanced binding and phagocytosis | (6) |
|
| Campylobacter jejuni | Sialoadhesin/Siglec-1 Siglec-7 | Sialic acids on LPS | Modulation of factors affecting helper T-cell differentiation | (7-9) |
|
| Group B Streptococcus type III | Siglec-9 | Sialic acids on LPS | Attenuated immune responses | (10) |
|
| Pseudomonas aeruginosa | Siglec-9 | Sialic acids on glycoproteins adsorbed from human bodily fluid | Attenuated immune responses | (11) |
|
| Group B Streptococcus type Ia | Siglec-5 Siglec-13 (chimpanzee) | β protein (independent of sialic acid) | Attenuated immune responses | (13, 23) |
|
| Eukaryotes | |||||
| Trypanosoma cruzi | Siglec-E (mouse) | Sialic acids acquired from human glycoproteins by trans-sialidase | Attenuated immune responses | (14) |
|
| Candida albicans | Siglec-7 | zymosan (?) | Enhanced immune responses | (25) |
|

Fig.2 How pathogens engage inhibitory Siglec, and how activating-type Siglec may counteract
Three different ways by which various pathogens engage inhibitory Siglec have been found: (I) synthesize sialic acids (Sias) on their own to interact with Siglecs; (II) borrow Sias from the host to interact with Siglecs; and (III) express protein that directly interacts with Siglecs. These interactions generally result in attenuated immune responses to the pathogens, to their advantage. Activating-type Siglec may counter these pathogens by recognizing them and enhancing immune responses against them.
Another case of "molecular mimicry of the host" employing Sia is found in Trypanosoma cruzi, a eukaryotic pathogen of a zoonotic disease called Chagas disease in Latin America. T. cruzi has an enzyme called trans-sialidase, which transfers Sias from the host glycoconjugates to the mucin-like glycoconjugates on its own cell surface. It has been postulated that the Sias on the pathogen recruit factor H and function as camouflage from the complement system, while a new study has proposed that Sia-dependent interaction with an inhibitory Siglec also provides a survival advantage to the pathogens by dampening the immune responses against them 14.
In addition to these cellular organisms, some viruses have also been reported to interact with Siglecs. The first example of a virus that interacts with a Siglec is the porcine reproductive and respiratory syndrome virus (PRRSV), which is a major threat to the swine industry. It was reported that Sias on the viral particles are necessary for the interaction with porcine sialoadhesin/Siglec-1 on macrophages, and this interaction enhances macrophage infection by the virus 15.
Human immunodeficiency virus (HIV) has also been reported to interact with human sialoadhesin/Siglec-1 to enhance binding to macrophages 16 and monocytes 17 (which aberrantly express sialoadhesin/Siglec-1 in the presence of interferons). Such interactions result either in the enhanced infection of macrophages 16 or in the enhanced trans-infection of T cells 17. These findings appear at odds with previous reports that gp120, the major envelope glycoprotein of HIV, is heavily glycosylated predominantly with high mannose-type N-glycans and interacts with C-type lectins expressed on the innate immune system, such as mannose-binding lectins (MBL) and dendritic cell-specific intercellular adhesion molecule-3-grabbing non-integrin (DC-SIGN) 18. However, gp120 is reported to carry sialylated complex-type N-glycans as well 19.
Other examples of viruses that interact with a Siglec include varicella zoster virus (VZV) and herpes simplex virus (HSV), both of which belong to herpesviridae and are closely related. These viruses remain dormant within sensory neurons (i.e., latent infection) after the initial infection, and their interaction with myelin-associated glycoprotein (MAG)/Siglec-4, which is expressed on the Schwann cells (in the peripheral nervous system) and oligodendrocytes (in the central nervous system) that insulate neurons, may be relevant to this viral tropism 20. It has been reported that viral glycoprotein B interacts with MAG/Siglec-4, but the involvement of Sias in the interaction was not explicitly demonstrated.
Given that the virally encoded envelope proteins undergo glycosylation by the host cell's Golgi apparatus, it seems natural that enveloped viruses carry Sias and interact with some Siglecs, at least in vitro. Thus, the number of examples of viruses interacting with Siglecs is expected to expand in the future.
In the examples explained so far, Siglecs are being exploited by pathogens to their advantage. Could there be a way to counter these exploitations? One possibility to counteract the threats is to set a "trap" against these pathogens, such as a Siglec that interacts with the pathogens but triggers pro-inflammatory, rather than anti-inflammatory, responses (Fig. 2). Among the small number of Siglecs with activating signaling properties, Siglec-14 appears to fit the bill. Siglec-14 has an extracellular domain that is extremely similar to that of Siglec-5, recognizes a similar set of sialylated glycan structures as Siglec-5, and has pro-inflammatory properties unlike Siglec-5, which has an inhibitory property. Moreover, genes encoding for Siglec-5 and Siglec-14 maintain unusually close similarity in multiple primate lineages 21. Many other pairs of immune receptors that show extreme sequence similarity with each other and have opposing signaling properties have been identified; it has been hypothesized that the activating-type receptor of the pair originates as a counter-measure against some pathogens that exploit its inhibitory counterpart 22.
However, it may be too simplistic to assume that the evolution of Siglecs always follows this pattern. A recent paper has suggested that Siglec-13, an activating-type Siglec lost in humans but present in other primates, interacts with group B Streptococcus type Ia in a Sia-independent (and β protein-dependent) manner but attenuates, rather than enhances, pro-inflammatory response 23. This may suggest that the effectiveness of activating-type Siglecs as a counter-measure against pathogens may not last very long. Since weak clustering of activating-type receptors sometimes leads to inhibitory signals (because the sequence motif involved in the recruitment of kinases to the activating-type receptors is similar to that involved in the recruitment of phosphatases to the inhibitory-type receptors) 24, the pathogens may fine-tune the density of the Siglec ligands so that the engagement with activating-type Siglec will not harm them. My colleagues and I have noted that the evolutionary "shelf life" of the activating-type Siglecs in general appears to be much shorter than that of inhibitory-type Siglecs, and many of them are lost in the process of evolution - they are either converted to pseudogenes or deleted from the genome altogether. In addition, another study reports that Siglec-7, which should have a negative regulatory property, binds zymosan (an yeast polysaccharide that contains no Sias) and elicits pro-inflammatory responses in monocytes 25. Thus, the interactions between pathogens and Siglecs and their consequences may be more complex than we have initially imagined.
The interaction between pathogens (or more broadly, microbes including pathogens, commensals, and probionts) and endogenous lectins is a fascinating area of research, which is sure to produce more insights into the fundamental biology of glycans and lectins in the future. Genetic polymorphisms of human Siglecs, some of which are null alleles resulting in the loss of some Siglecs (e.g., Siglec-12, -14, and -16; refs 26-28), may have some relevance to their interaction with some pathogens. Although more stringent validations may be needed in some cases to demonstrate that these interactions are actually at work in vivo (e.g., by the laboratory studies using animal models and/or by the epidemiological studies analyzing relevant human polymorphisms), these interactions can be potential targets of therapeutic interventions. Thus, interactions between Siglecs and various microbes deserve more attention.
Acknowledgment
Professor Ajit Varki of the University of California, San Diego provided many helpful comments and suggestions for which I am very grateful.



