
Reiji Kannagi
Graduated Kyoto University School of Medicine in 1974 and obtained his Ph.D. from Kyoto University Graduate School of Medicine in 1982. Started his professional career in the laboratory of Prof. Sen-itiroh Hakomori at Fred Hutchinson Cancer Research Center, Program of Biochemical Oncology, Seattle, Washington, USA (1980-1982). He was assistant professor (1982-1985) then lecturer (1985-1991) at Kyoto University School of Medicine. He is presently chief of the Department of Molecular Pathology at Aichi Cancer Center Research Institute. He received the Young Investigator Award from the Japanese Biochemical Society in 1985, and from the Japanese Association for Cancer Research in 1986. His research interests include cell-to-cell interaction and recognition, biochemistry of carbohydrates, and medical oncology.
Selectin is a family of cell adhesion molecules which mediate the carbohydrate-protein interaction with specific ligands and interrupt the rapid flow of leukocytes in the circulation by initiation of rolling and tethering on endothelial cells. Cell adhesion mediated by selectin is implicated in extravasation of leukocytes, homing of lymphocytes, and infiltration or metastasis of malignant cells including leukemia and cancer cells. The family consists of three members, E-, P- and L-selectins (Fig. 1). Each selectin has a calcium-dependent carbohydrate-binding domain (C-type lectin domain) in its amino terminus, followed by an epidermal growth factor (EGF)-like domain, a varying number of complement-binding protein-like repeats, a transmembrane domain and a short cytoplasmic domain. Selectin-mediated cell adhesion is known to be partly regulated by the expression level of selectin molecules per se. It is increasingly clear, however, that selectin-mediated cell adhesion is also regulated dynamically by the expression of specific carbohydrate ligands, which are recognized by the C-type lectin domain in the selectin molecules. Because the C-type lectin domains of the three members of the selectin family are considerably homologous, it has been predicted that the three members of the selectin family have similar ligand binding specificities. In fact, each member of the family has distinct carbohydrate ligand specificity, which governs the complex processes of specific homing of various subsets of leukocytes and inflammatory mobilization of activated leukocytes.1
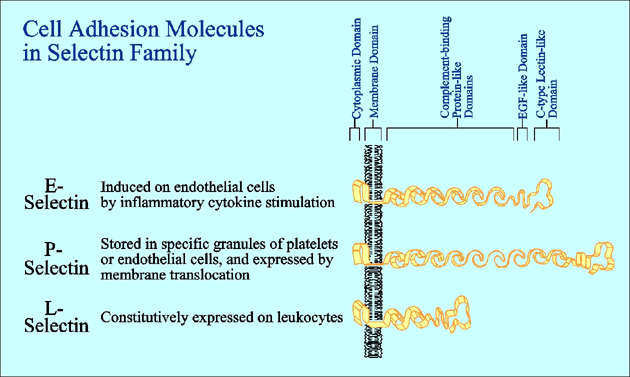
Fig. 1 Schematic illustration of three members of the selectin family
E-selectin (ELAM-1, or CD62E) is known to be inducibly expressed on endothelial cells at the site of inflammation through the action of inflammatory cytokines such as IL-1 or TNFα, and thought to be involved in the recruitment of leukocytes during the course of inflammation. In vitro stimulation with these inflammatory cytokines is known to induce transcription of E-selectin mRNA in cultured endothelial cells mediated by multiple binding sites for NF-κB-like transcriptional factors located in the 5'-regulatory region of the E-selectin gene. Maximum E-selectin protein expression at the cell surface is attained within 4 hours, and a significant decrease in its expression is apparent 12 ~ 24 hours after the stimulation. The carbohydrate ligand for E-selectin on leukocytes was shown to be sialyl Lewisx in the early 1990’s.2-4 The E-selectin-mediated leukocyte endothelial cell interaction is mediated by the binding of the C-type lectin-like domain of E-selectin to the sialyl Lewisx determinant expressed on leukocytes. Rolling of leukocytes mediated by E-selectin and its specific ligands is shown in Video 1.
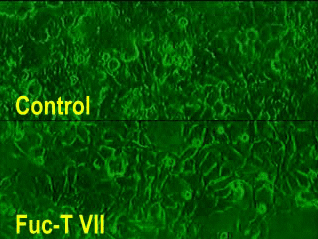
Video 1 (click the Figure)
An example of rolling assay using E-selectin expressing cells and the cells transfected with Fuc-T VII gene (lower panel, labeled as Fuc-T VII). Upper panel shows the rolling with the negative control cells (upper panel, labeled as Control). Note that only the lower panel shows significant rolling of E-selectin-expressing cells. Both panels recorded at the shear force of 1.13 dyne/cm2.
Sialyl Lewisx is also frequently expressed on epithelial cancer cells, and regarded as a cancer-associated carbohydrate determinant. Another well-known cancer-associated carbohydrate determinant, sialyl Lewisa, also serves as a ligand for E-selectin (Fig. 2).5,6 Cancer cells expressing sialyl Lewisx or sialyl Lewisa strongly adhere to activated endothelial cells in the monolayer cell adhesion assay. The sialyl Lewisx and sialyl Lewisa determinants expressed on cancer cells are thought to be involved in the process of hematogenous metastasis of cancers.7
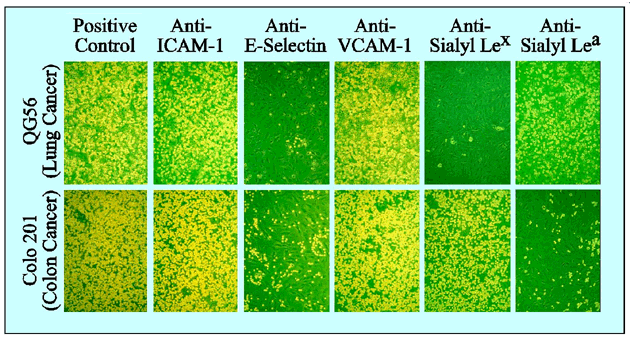
Fig. 2
E-selectin-mediated adhesion of human cancer cells to endothelial cells Typical examples of non-static monolayer cell adhesion experiments indicating adhesion of cultured human cancer cells to IL-1β-stimulated human umbilical vein endothelial cells. Note that adhesion of a cultured human lung cancer cell line QG56 is inhibited by anti-sialyl Lewisx antibody, while that of cultured human colon cancer cell line COLO201 is inhibited by anti-sialyl Lewisa antibody. Adhesion of both lines is completely inhibited by anti-E-selectin antibody.
There is considerable evidence showing that carbohydrate ligands for E-selectin are not limited to sialyl Lewisx and sialyl Lewisa. Some internally fucosylated structures are reportedly capable of binding E-selectin, even when the determinant does not carry a terminal sialyl Lewisx/a structure.8-10 E-selectin ligands on the cell surface are a set of heterogenous sialyl Lewisx/a-like determinants.
Sialyl Lewisx on leukocytes has been reported to be carried by multiple glycoproteins. In earlier studies, L-selectin was proposed to be one of the major core proteins that carry carbohydrate ligands for E-selectin.11 PSGL-1, originally described as a specific core protein for P-selectin, provides sialyl Lewisx determinants also to E-selectin for binding. A unique 150 kDa core protein with N-linked carbohydrate side chains, ESL-1 (E-selectin ligand-1), is also reported to occur in murine myeloid cells.12 In cancer cells, sialyl Lewisx and sialyl Lewisa determinants are known to be carried by high-molecular-weight mucin molecules, such as MUC-1, as well as by glycolipids. Most researchers agree that E-selectin preferentially recognizes the carbohydrate portion of the ligand molecules, and does not require a specific core protein for its binding, while core proteins would indirectly affect the binding activity through optimizing the density and conformation of the carbohydrate ligands at the cell surface.
P-selectin (PADGEM, GMP-140, or CD62P) is expressed on endothelial cells and activated platelets, and reacts with the carbohydrate ligands expressed on leukocytes. The physiological significance of P-selectin was originally thought to be the leukocyte-platelet interaction and leukocyte-endothelial adhesion in the inflammatory processes. P-selectin is known to be stored in α granules in platelets and Wibel-Palade granules in endothelial cells, and is rapidly expressed by membrane translocation upon stimulation. P-selectin is also known to be inducibly expressed in murine endothelial cells through relatively slower kinetics similar to human E-selectin.
Earlier studies demonstrated that recombinant P-selectin binds to sialyl Lewisx and sialyl Lewisa.13This was readily predicted since these carbohydrate determinants had earlier been shown to serve as ligands for E-selectin, and the C-type lectin domains of E- and P-selectins are highly homologous to each other. There are, however, several significant differences between the E-selectin and P-selectin ligands. In binding to immobilized sialyl Lewisx, recombinant P-selectin characteristically requires a 5- to 10-times higher concentration to attain the binding level obtained with E-selectin. P-selectin-mediated cell adhesion is known to be abrogated by inhibitors of sulfation such as NaClO3, whereas E-selectin-mediated cell adhesion is not. The P-selectin ligands on leukocytes were known to be protease-sensitive, whereas those for E-selectin were shown to be quite resistant to protease treatment. These findings eventually led to the discovery of PSGL-1 (P-selectin glycoprotein ligand-1), a 220 kDa membrane glycoprotein expressed on most leukocytes. PSGL-1 should carry a sialyl Lewisx determinant on its carbohydrate side chains, and should be sulfated at the tyrosine residues near its amino terminus for optimum binding to P-selectin.14-16 The sialyl Lewisx determinant and sulfated tyrosine residue in PSGL-1 peptide have both been proven necessary for P-selectin to bind cells, while E-selectin requires only sialyl Lewisx determinant. The dimeric form of PSGL-1 was also found necessary to interact with P-selectin. The binding affinity of P-selectin to PSGL-1 properly modified by sialyl Lewisx determinant is very high; the Km value reportedly reaches around 10-9 M. Thus, PSGL-1 is so far the clearest example of specifically required core proteins for selectin ligands ever reported, although the exact role of the sulfated tyrosine residue is still ambiguous in that P-selectin does not seem to discriminate which among the three candidate tyrosine residues in the N-terminal peptide sequence of PSGL-1 is sulfated .
L-selectin (LAM-1, LECAM-1, gp90mel, or CD62L) is constitutively expressed on leukocytes. L-selectin was first described as a lymph node receptor involved in the homing of naïve T lymphocytes to peripheral lymph nodes.17,18 High endothelial venules (HEV) are known to serve as an entrance for those lymphocytes homing into peripheral lymph nodes. Since L-selectin is expressed on leukocytes, the HEV endothelial cells should express the corresponding carbohydrate ligand for L-selectin, which earlier studies have suggested to be sialyl Lewisx.
However, attempts to identify the L-selectin ligand on high endothelial cells using monoclonal anti-sialyl Lewisx antibodies resulted in complicated results.19 The high endothelial cells in human lymph nodes were stained by only a limited group of anti-sialyl Lewisx antibodies, but not by the other group of anti-sialyl Lewisx antibodies.19,20 The antibodies that stained high endothelial cells also inhibited the L-selectin-dependent cell adhesion to HEV endothelial cells.19 These findings indicated that a unique sialyl Lewisx-like determinant, but not the authentic sialyl Lewisx, was present on high endothelial cells, and served as a ligand for L-selectin. This was further supported by the finding that most anti-Lewisx antibodies failed to stain sialidase-treated HEV endothelial cells. The genuine sialyl Lewisx, after sialidase treatment, should have yielded conventional Lewisx, which would have been readily detected by usual anti-Lewisx antibodies.
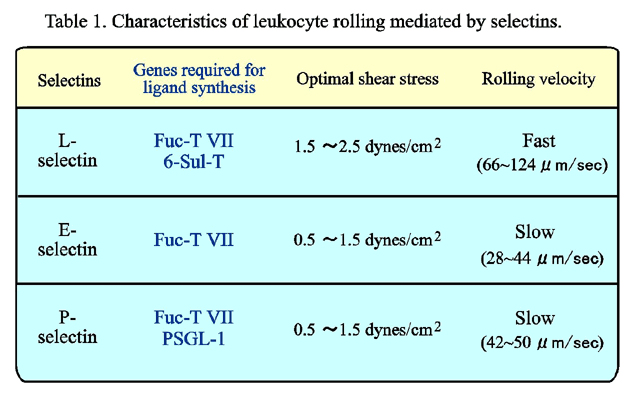
This riddle was solved by the recent discovery of sulfated sialyl Lewisx determinants on HEV, as the modification by sulfate blocked the binding of most of the anti-sialyl Lewisx and anti-Lewisx antibodies, while the binding of a few other antibodies was not affected by sulfation of the sialyl Lewisx determinants. The carbohydrate ligand for L-selectin expressed on HEV in human peripheral lymph nodes was identified as sialyl 6-sulfo Lewisx,21 while 6'-sulfated and 6,6'-disulfated structures were proposed to be predominant for HEV in murine peripheral lymph nodes.22,23 Sialyl 6-sulfo Lewisx is expressed on HEV in peripheral lymph nodes, as well as HEV-like endothelial cells in Peyer's patches, appendices and so called gut-associated lymphoreticular tissues (GALT) (Fig. 3). Considerable difference in the carbohydrate ligands for L-selectin is found between humans and mice, but 6-sulfation seems to be essential in both species, as functional L-selectin ligands can be reconstituted by the transfection of co-transfection of a fucosyltransferase gene and a 6-sulfotransferase gene.24 The L-selectin ligand thus reconstituted mediated significant rolling of L-selectin-expressing cells with a rolling velocity of around 60~120 µm/sec,25 a range comparable to that previously reported for L-selectin-mediated rolling in vivo.26 Two genes for 6-sulfotransferase isoenzymes, the candidate enzymes for the synthesis of sialyl 6-sulfo Lewisx in HEV, were cloned,27,28 and the disruption of one of the genes in mice was shown to reduce L-selectin-mediated lymphocyte homing to 50 % of that in wild-type mice.29
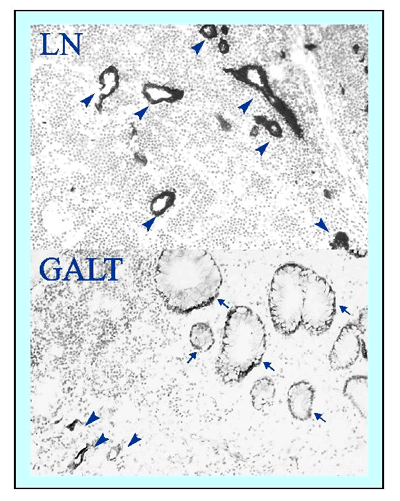
Fig.3
Distribution of sialyl 6-sulfo Lewisx, the specific carbohydrate ligand for L-selectin, in high endothelial venules of peripheral lymph node (upper panel) and gut-associated lymphoreticular tissue (lower panel) Results of immunohistochemical staining using specific antibody G152. Arrowheads indicate high endothelial venule-like blood vessels expressing sialyl 6-sulfo Lewisx, and thin arrows indicate intestinal epithelial cells, which also express the determinant.
Several glycoproteins on HEV endothelial cells are proposed as L-selectin ligand-specific core proteins, which carry the carbohydrate ligands and provide them to L-selectin. These include GlyCAM-1, CD34, Sgp150/>200, and po docalyxin.30-33 PSGL-1 was proposed to facilitate high affinity binding of L-selectin.34-37 Since endothelial cells generally lack expression of PSGL-1, this must be limited to the L-selectin mediated leukocyte-leukocyte interaction, and may not be involved in the L-selectin mediated leukocyte-endothelial cell interaction. As yet there is no definitive evidence to support an absolute requirement for a specific core protein for L-selectin ligand activity. GlyCAM-1 has not been described for humans yet. Impairment of lymphocyte homing is not prominent in CD34 knockout mice.38 The cells that express none of these core proteins but transfected with only Fuc-T VII and 6-sulfotransferase genes acquire sufficient ability to support the rolling of L-selectin-expressing cells at a velocity comparable to physiological L-selectin-mediated rolling. These findings imply that the carbohydrate portion of the L-selectin ligand plays the essential role, while again core proteins would indirectly affect the binding activity through optimizing the density and conformation of the carbohydrate ligand at the cell surface.
Earlier results of ELISA employing recombinant human selectin-Ig indicated that all selectins are capable of binding to sialyl Lewisx determinant.13 Sialyl 6-sulfo Lewisx determinant was also reported to react with all selectins.39 However, ELISA frequently picks up low-affinity bindings that are not always physiologically relevant. We tried cell-binding studies employing the lymphoid cells expressing both sialyl 6-sulfo Lewisx and conventional sialyl Lewisx, and also endogenous PSGL-1.40 Binding of E- and P-selectins to these cells was partially inhibited by anti-sialyl Lewisx antibody, or by anti-sialyl 6-sulfo Lewisx antibody (Fig. 4). Complete inhibition was observed only when the mixture of the two antibodies was used in the inhibition experiments. On the other hand, binding of L-selectin to these cells was barely affected by anti-sialyl Lewisx antibody, but was completely inhibited by anti-sialyl 6-sulfo Lewisx antibody (Fig. 4).40 These results indicate that both sialyl 6-sulfo Lewisx and conventional sialyl Lewisx can serve as ligands for E- and P-selectins, while L-selectin is quite specific to sialyl 6-sulfo Lewisx (Fig. 5). Anti-PSGL-1 antibody treatment almost completely abrogated the binding of P-selectin, yet only partially affected the binding of L-selectin, and barely affected the binding of E-selectin (Fig. 4).40 This confirmed the well-established importance of PSGL-1 in P-selectin-mediated adhesion (Fig. 5). These results suggest the possibility, however, that PSGL-1 may not exert a facilitative effect on L-selectin-mediated adhesion as was anticipated.
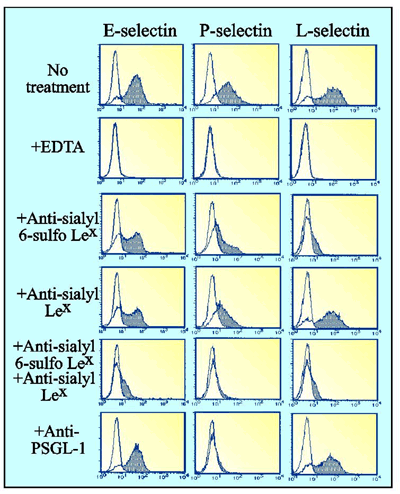
Fig.4
Contribution of carbohydrate determinants and core protein PSGL-1 in the binding of E-, P- and L-selectin to human lymphoid cells expressing both sialyl Lewisx and sialyl 6-sulfo Lewisx Results of flow cytometric analysis of binding of recombinant selectin-Ig are shown.40
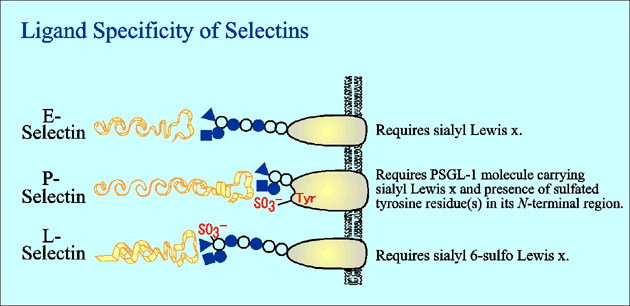
Fig.5 Schematic illustration of specific ligands for three members of the selectin family
We recently generated COS7 cells transfected with the genes encoding PSGL-1, 6-Sul-T and Fuc-T VII, and tested them in rolling assays under shear flow.41 The triple-transfected cells fully supported rolling mediated by all three selectins, and provided the best experimental system to compare the ligand specificity of each selectin. We carefully compared the adhesive behavior of these fully potent cells with several other transfectant cells that were generated by omitting each one of these three genes, or by replacing it with non-functional mutant genes. L-selectin-mediated rolling was virtually unaffected by the replacement of wild-type PSGL-1 in fully potent cells with a mutant lacking three critical tyrosine residues at its N-terminus. L-selectin-mediated adhesion was also virtually unaffected by the treatment of the fully potent cells with mocarhagin, which cleaved a short peptide containing sulfated tyrosine residues from PSGL-1. In contrast, L-selectin-mediated rolling was drastically impaired and reduced to only 6 % of that of fully potent cells, when the transfection of 6-sulfotransferase gene was omitted and the cells expressed only PSGL-1 and Fuc-T VII genes.41
On the other hand, P-selectin-mediated rolling was completely eliminated when PSGL-1 was either mutated or cleaved by mocarhagin, whereas the cells expressing PSGL-1 and Fuc-T VII but not 6-sulfotransferase genes showed only a modest decrease in P-selectin-mediated adhesion compared to the fully potent cells.41 E-selectin-mediated adhesion was sulfation-independent, requiring only Fuc-T VII. These results,41 together with our previous findings,40 clearly established that the three members of the selectin family have distinct requirements for ligand sulfation, as summarized in Table 1.
Proteins and nucleic acids are generally believed to be indispensable functional molecules, but the role of carbohydrates as the third functional molecules has long remained obscure, and great physiological significance has been attributed to aglycones such as core proteins rather than to carbohydrate portions. Recent progress, however, indicates that the carbohydrate portion of selectin ligands plays the key role in the regulation of selectin-mediated cell adhesion. This may herald the advent of a new age for functional glycomics.
Granulocytes and monocytes constitutively express carbohydrate ligands for selectins. In contrast, carbohydrate ligands on lymphocytes are highly inducible. Selectin ligands on leukocytes are mainly synthesized by Fuc-T VII and Fuc-T IV. Most resting lymphocytes in human peripheral blood do not express sialyl Lewisx, but its expression is strongly induced when lymphocytes are activated by inflammatory stimuli, and activated lymphocytes exhibit vigorous adhesion to endothelial P- and E-selectins. This accompanies remarkable induction of Fuc-T VII and IV gene transcription. Dynamic regulation of transcription of Fuc-T VII and IV genes is observed upon stimulation of leukocytes by inflammatory cytokines.42-48
On the other hand, PSGL-1, the most important core protein for selectin ligands on leukocytes, is constitutively expressed by a wide variety of human leukocytes. The cells that express sialyl Lewisx almost always express PSGL-1, but the reverse is not necessarily true. Expression of PSGL-1 does not show drastic change under inflammatory stimuli. These findings suggest that the acquisition of cellular selectin binding activity upon inflamatory stimulation is achieved mainly by the induction of sialyl Lewisx expression by transcriptional activation of fucosyltransferase genes, rather than by the inducible expression of PSGL-1 peptide synthesis. Several candidates for protein O-sulfotransferases that sulfate tyrosine moieties in PSGL-1 were recently identified,49,50 but it remains unknown if these enzymes are actively involved in the regulation of P-selectin-mediated cell adhesion.
Some researchers suggest that L-selectin may also be implicated in the leukocyte-leukocyte interaction and/or neutrophil-endothelial interaction at the site of inflammation.35,51 If L-selectin is involved in the adhesion of leukocytes to vascular endothelial cells at the site of an inflammatory lesion, the putative L-selectin ligand on endothelial cells should be inducibly expressed by the inflammatory stimuli. Regulation of 6-sulfotransferases remains largely unknown at this juncture, but at least a moderate induction of its transcription was noted upon stimulation by inflammatory cytokines.24
Another example of the vital role of carbohydrate ligands in the regulation of selectin-mediated cell adhesion is the rapid inactivation of selectin binding activity by post-translational modification of the sialic acid moiety in carbohydrate ligands for selectins. Sialic acid residue is an indispensable component in carbohydrate ligands for selectins. When stimulation of the cells triggers mobilization of calcium ion, the sialic acid moiety is converted to cyclic sialic acid, and the ligand loses the binding activity to selectins. This was first observed in the sialyl 6-sulfo Lewisx carried on human lymphoid cells,52 and also later found on some cells of epithelial origin.53 The conversion of active selectin ligands to inert ligands is a very rapid process that is completed within 5 minutes when cells are stimulated by calcium ionophore, and mediated by the enzyme tentatively termed sialic acid cyclase.43,54,55 This kind of rapid modulation of cell surface carbohydrates that is accompanied by a physiologically significant functional consequence (e.g., negative feedback of cell adhesive activity in this example), has only recently come to light, and indicates that cell surface carbohydrates are well qualified to support vital physiological functions.



