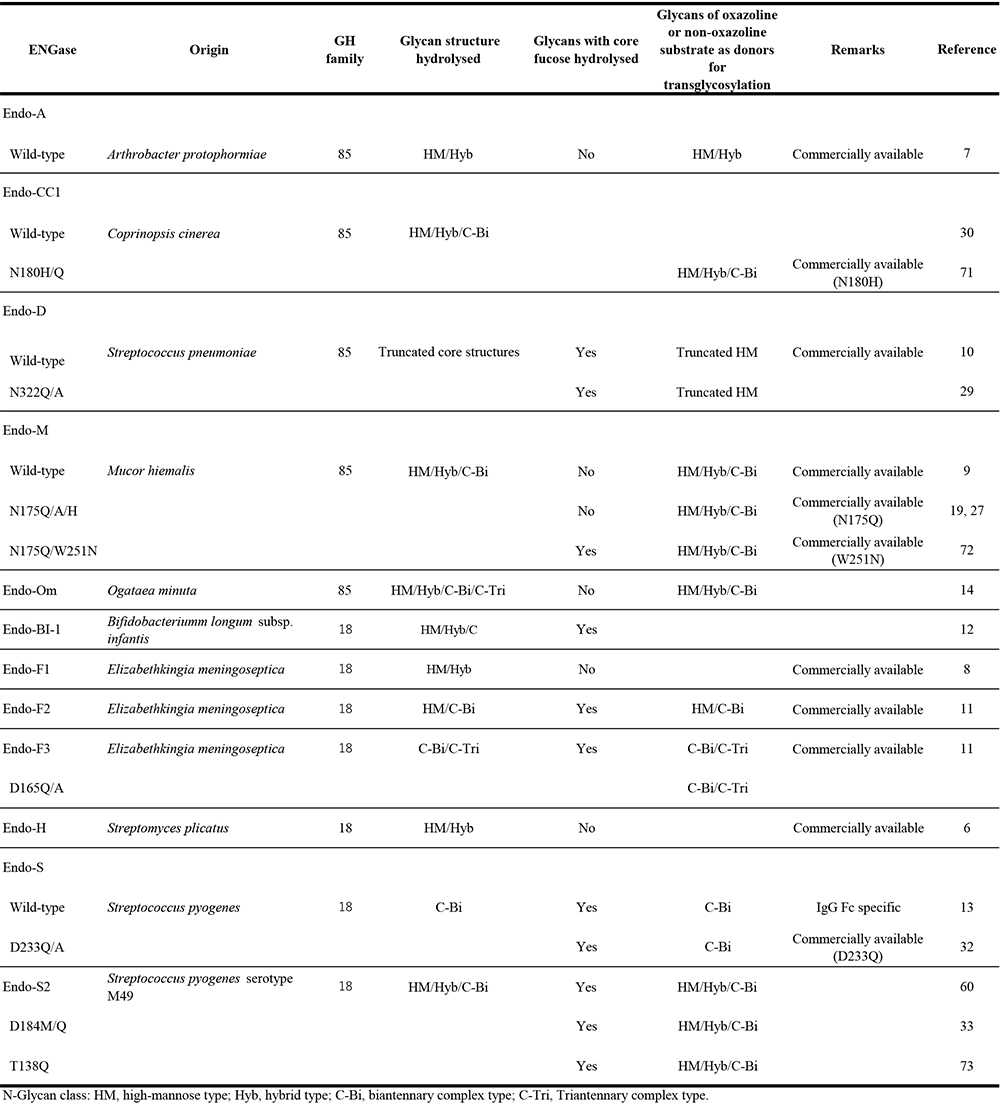
Table 1. Various useful microbial ENGases for the glycoengineering
一次構造に基づいてENGaseには二つのタイプがあり、一つはグリコシドヒドロラーゼ(GH)ファミリー18として分類され、他方はCarbohydrate Active enZymes(CAZy)データベースにおいてGHファミリー85として分類されている。全てのGHファミリー18の酵素は活性基に2つのカルボン酸残基、すなわちグルタミン酸とアスパラギン酸を有している一方、GH85ファミリーの酵素は1残基のカルボン酸残基、すなわちグルタミン酸しか有していない。もう一方の重要な触媒残基はアスパラギンである。全てのENGaseは8つのα-ヘリックスと8つの平行なβ-シート構造からなる同じ(β/α)8-TIM バレル構造を有する。同じ酵素活性を示しながら、GHファミリー内のほとんどの酵素は25~30%の同一配列しか示さず、これらの酵素の間には、異なる触媒機構と異なる基質特異性を示す多くの違いがあることを示唆している。
Wong C.-H., Haynie S.L., Whitesides G.M. Enzyme-catalyzed synthesis of N-acetyllactosamine with in situ regeneration of uridine 5'-diphosphate glucose and uridine 5'-diphosphate galactose. J. Org. Chem. 1982, 47:5416–5418.
Fairbanks A.J. The ENGases: versatile biocatalysts for the production of homogeneous N-linked glycopeptides and glycoproteins. Chem. Soc. Rev. 2017, 46:5128–5146.
Kobata A. Use of endo- and exoglycosidases for structural studies of glycoconjugates. Anal. Biochem. 1979, 100:1–14.
Karav S., Casaburi G., Arslan A., Kaplan M., Sucu B., Frese S. N-glycans from human milk glycoproteins are selectively released by an infant gut symbiont in vivo. J. Funct. Foods. 2019, 61:103485.
Sastre D.E., Sultana N., Navarro M.V.A.S., Huliciak M., Du J., Cifuente J.O., Flowers M., Liu X., Lollar P., Trastoy B., Guerin M.E., Sundberg E.J. Human gut microbes express functionally distinct endoglycosidases to metabolize the same N-glycan substrate. Nat. Commun. 2024, 15:5123.
Tarentino A.L., Plummer T.H., Maley F. A re-evaluation of the oligosaccharide sequence associated with ovalbumin. J. Biol. Chem. 1972, 247:2629–2631.
Takegawa K., Nakoshi M., Iwahara S., Yamamoto K., Tochikura T. Induction and purification of endo-beta-N-acetylglucosaminidase from Arthrobacter protophormiae grown in ovalbumin. Appl. Environ. Microbiol. 1989, 55:3107–3112.
Elder J.H., Alexander S. Endo-beta-N-acetylglucosaminidase F: endoglycosidase from Flavobacterium meningosepticum that cleaves both high-mannose and complex glycoproteins. Proc. Natl. Acad. Sci. U.S.A. 1982, 79:4540–4544.
Kadowaki S., Yamamoto K., Fujisaki M., Izumi K., Tochikura T., Yokoyama T. Purification and characterization of a novel fungal endo-beta-N-acetylglucosaminidase acting on complex oligosaccharides of glycoproteins. Agric. Biol. Chem. 1990, 54:97–106.
Muramatsu T. Demonstration of an endo-glycosidase acting on a glycoprotein. J. Biol. Chem. 1971, 246:5535–5537.
Huang W., Li J., Wang L.-X. Unusual transglycosylation activity of Flavobacterium meningosepticum endoglycosidases enables convergent chemoenzymatic synthesis of core fucosylated complex N-glycopeptides.
ChemBioChem. 2011, 12:932–941.
Garrido D., Nwosu C., Ruiz-Moyano S., Aldredge D., German J.B., Lebrilla C.B., Mills D.A. Endo-b-N-acetylglucosaminidases from infant gut-associated Bifidobacteria release complex N-glycans from human milk glycoproteins. Mol. Cell. Proteomics. 2012, 11:775–785.
Collin M., Olsén A. EndoS, a novel secreted protein from Streptococcus pyogenes with endoglycosidase activity on human IgG. EMBO J. 2001, 20:3046–3055.
Murakami S., Takaoka Y., Ashida H., Yamamoto K., Narimatsu H., Chiba Y. Identification and characterization of endo-β-N-acetylglucosaminidase from methylotrophic yeast Ogataea minuta. Glycobiology. 2013, 23:736–744.
Edelman J. The formation of oligosaccharides by enzymic transglycosylation. Adv. Enzymol. Relat. Subj. Biochem. 1956, 17:189–232.
Rich J.R., Withers S.G. Emerging methods for the production of homogeneous human glycoproteins. Nat. Chem. Biol. 2009, 5:206–215.
Wang L.-X. Chemoenzymatic synthesis of glycopeptides and glycoproteins through endoglycosidase-catalyzed transglycosylation. Carbohydr. Res. 2008, 343:1509–1522.
Yamamoto K. Recent advances in glycotechnology for glycoconjugate synthesis using microbial endoglycosidases. Biotech. Lett. 2013, 35:1733–1743.
Umekawa M., Huang W., Li B., Fujita K., Ashida H., Wang L.-X., Yamamoto K. Mutants of Mucor hiemalis endo-beta-N-acetylglucosaminidase show enhanced transglycosylation and glycosynthase-like activities. J. Biol. Chem. 2008, 283:4469–4479.
Mackenzie L.F., Wang Q., Warren R.A.J., Withers S.G. Glycosynthases: mutant glycosidases for oligosaccharide synthesis. J. Am. Chem. Soc. 1998, 120:5583–5584.
Alsina C., Faijes M., Planas A. Glycosynthase-type GH18 mutant chitinases at the assisting catalytic residue for polymerization of chitooligosaccharides. Carbohydr. Res. 2019, 478:1–9.
Koshland D.E. Stereochemistry and the mechanism of enzymatic reactions. Biol. Rev. 1953, 28:416–436.
Mark B.L., Vocadlo D.J., Knapp S., Triggs-Raine B.L., Withers S.G., James M.N.G. Crystallographic evidence for substrate-assisted catalysis in a bacterial beta-hexosaminidase. J. Biol. Chem. 2001, 276:10330–10337.
Fujita M., Shoda S., Haneda K., Inazu T., Takegawa K., Yamamoto K. A novel disaccharide substrate having 1,2-oxazoline moiety for detection of transglycosylating activity of endoglycosidases. Biochim. Biophys. Acta. 2001,1528:9–14.
Cobucci-Ponzano B., Strazzulli A., Rossi M., Moracci M. Glycosynthases in biocatalysis. Adv. Synth. Catal. 2011, 353:2284–2300.
Umekawa M., Li C., Higashiyama T., Huang W., Ashida H., Yamamoto K., Wang L.-X. Efficient glycosynthase mutant derived from Mucor hiemalis endo-beta-N-acetylglucosaminidase capable of transferring oligosaccharide from both sugar oxazoline and natural N-glycan. J. Biol. Chem. 2010, 285:511–521.
Huang W., Li C., Li B., Umekawa M., Yamamoto K., Zhang X., Wang L.-X. Glycosynthases enable a highly efficient chemoenzymatic synthesis of N-glycoproteins carrying intact natural N-glycans. J. Am. Chem. Soc. 2009, 131:2214–2223.
Fan S.-Q., Huang W., Wang L.-X. Remarkable transglycosylation activity of glycosynthase mutants of endo-D, an endo-β-N-acetylglucosaminidase from Streptococcus pneumoniae. J. Biol. Chem. 2012, 287:11272–11281.
Higuchi Y., Eshima Y., Huang Y., Kinoshita T., Sumiyoshi W., Nakakita S., Takegawa K. Highly efficient transglycosylation of sialo-complex-type oligosaccharide using Coprinopsis cinerea endoglycosidase and sugar oxazoline. Biotech. Lett. 2017, 39:157–162.
Goodfellow J.J., Baruah K., Yamamoto K., Bonomelli C., Krishna B., Harvey D.J., Crispin M., Scanlan C.N., Davis B.G. An endoglycosidase with alternative glycan specificity allows broadened glycoprotein remodelling. J. Am. Chem. Soc. 2012, 134:8030–8033.
Huang W., Giddens J., Fan S.-Q., Toonstra C., Wang L.-X. Chemoenzymatic glycoengineering of intact IgG antibodies for gain of functions. J. Am. Chem. Soc. 2012, 134:12308–12318.
Li T., Tong X., Yang Q., Giddens J.P., Wang L.-X. Glycosynthase mutants of endoglycosidase S2 show potent transglycosylation activity and remarkably relaxed substrate specificity for antibody glycosylation remodeling. J. Biol. Chem. 2016, 291:16508–16518.
Yamamoto K., Fujimori K., Haneda K., Mizuno M., Inazu T., Kumagai H. Chemoenzymatic synthesis of a novel glycopeptide using a microbial endoglycosidase. Carbohydr. Res. 1997, 305:415–422.
Mizuno M., Haneda K., Iguchi R., Muramoto I., Kawakami T., Aimoto S., Yamamoto K., Inazu T. Synthesis of a glycopeptide containing oligosaccharides: chemoenzymatic synthesis of eel calcitonin analogues having natural N-linked oligosaccharides. J. Am. Chem. Soc. 1999, 121:284–290.
Higashiyama T., Umekawa M., Nagao M., Katoh T., Ashida H., Yamamoto K. Chemo-enzymatic synthesis of the glucagon containing N-linked oligosaccharide and its characterization. Carbohydr. Res. 2018, 455:92–96.
Haneda K., Inazu T., Mizuno M., Iguchi R., Tanabe H., Fujimori K., Yamamoto K., Kumagai H., Tsumori K., Munekata E. Chemo-enzymatic synthesis of a bioactive peptide containing a glutamine-linked oligosaccharide and its characterization. Biochim. Biophys. Acta. 2001, 1526:242–248.
Saskiawan I., Mizuno M., Inazu T., Haneda K., Harashima S., Kumagai H., Yamamoto K. Chemo-enzymatic synthesis of the glycosylated alpha-mating factor of Saccharomyces cerevisiae and analysis of its biological activity. Arch. Biochem. Biophys. 2002, 406:127–134.
Umemura M., Itoh M., Makimura Y., Yamazaki K., Umekawa M., Masui A., Matahira Y., Shibata M., Ashida H., Yamamoto K. Design of a sialylglycopolymer with a chitosan backbone having efficient inhibitory activity against influenza virus infection. J. Med. Chem. 2008, 51:4496–4503.
Tomabechi Y., Suzuki R., Haneda K., Inazu T. Chemo-enzymatic synthesis of glycosylated insulin using a GlcNAc tag. Bioorg. Med. Chem. 2010, 18:1259–1264.
Wang L.-X., Song H., Liu S., Lu H., Jiang S., Ni J., Li H. Chemoenzymatic synthesis of HIV-1 gp41 glycopeptides: effects of glycosylation on the anti-HIV activity and α-helix bundle-forming ability of peptide C34. ChemBioChem. 2005, 6:1068–1074.
Li H., Singh S., Zeng Y., Song H., Wang L.‑X. Chemoenzymatic synthesis of CD52 glycoproteins carrying native N‑glycans. Bioorg. Med. Chem. Lett. 2005, 15:895–898.
Kowalczyk R., Brimble M.A., Tomabechi Y., Fairbanks A.J., Fletcher M., Hay D.L. Convergent chemoenzymatic synthesis of a library of glycosylated analogues of pramlintide: structure–activity relationships for amylin receptor agonism. Org. Biomol. Chem., 2014, 12:8142–8151.
Hojo H., Tanaka H., Hagiwara M., Asahina Y., Ueki A., Katayama H., Nakahara Y., Yoneshige A., Matsuda J., Ito Y., Nakahara Y. Chemoenzymatic synthesis of hydrophobic glycoprotein: synthesis of saposin c carrying complex-type carbohydrate. J. Org. Chem. 2012, 77:9437–9446.
Umekawa M., Higashiyama T., Koga Y., Tanaka T., Noguchi M., Kobayashi A., Shoda S., Huang W., Wang L.-X., Ashida H., Yamamoto K. Efficient transfer of sialo-oligosaccharide onto proteins by combined use of a glycosynthase-like mutant of Mucor hiemalis endoglycosidase and synthetic sialo-complex-type sugar oxazoline. Biochim. Biophys. Acta. 2010, 1800:1203–1209.
Kornfeld S. Structure and function of the mannose 6-phosphate/insulinlike growth factor II receptors. Annu. Rev. Biochem. 1992, 61: 307–330.
Chiba Y., Sakuraba H., Kotani M., Kase R., Kobayashi K., Takeuchi M., Ogasawara S., Maruyama Y., Nakajima T., Takaoka Y., Jigami Y. Production in yeast of alpha-galactosidase A, a lysosomal enzyme applicable to enzyme replacement therapy for Fabry disease. Glycobiology. 2002, 12:821–828.
Chiba Y., Akeboshi H. Glycan engineering and production of 'humanized' glycoprotein in yeast cells. Biol. Pharm. Bull. 2009, 32:786–795.
Priyanka P., Parsons T.B., Miller A., Platt F.M., Fairbanks A.J. Chemoenzymatic synthesis of a phosphorylated glycoprotein. Angew. Chem. Int. Ed. Engl. 2016, 55:5058–5061.
Yamaguchi T., Amin M.N., Toonstra C., Wang L.-X. Chemoenzymatic synthesis and receptor binding of mannose-6-phosphate (M6P)-containing glycoprotein ligands reveal unusual structural requirements for M6P receptor recognition. J. Am. Chem. Soc. 2016, 138:12472–12485.
Zhang X., Liu H., Meena N., Li C., Zong G., Raben N., Puertollano R., Wang L.-X. Chemoenzymatic glycan-selective remodeling of a therapeutic lysosomal enzyme with high-affinity M6P-glycan ligands. Enzyme substrate specificity is the name of the game. Chem. Sci. 2021, 12:12451–12462.
Shinoda C., Kitakaze K., Sasai Y., Nishioka S., Kobayashi I., Sumitani M., Tatematsu K., Iizuka T., Harazono A., Mitani A., Kaneko A., Imamura M., Miyabe-Nishiwaki T., Go Y., Hirata A., Takeuchi Y., Mizuno T., Kiriyama K., Tsukimoto J., Nadanaka S., Ishii-Watabe A., Kinoshita T., Kitagawa H., Suzuki Y., Oishi T., Sezutsu H., Itoh K. N-glycan-modified α-L-iduronidase produced by transgenic silkworms ameliorates clinical signs in a Japanese macaque with mucopolysaccharidosis I. Commun. Med. 2025, 5:128.
Beck A., Wurch T., Bailly C., Corvaia N. Strategies and challenges for the next generation of therapeutic antibodies. Nat. Rev. Immunol. 2010, 10:345–352.
Mizushima T., Yagi H., Takemoto E., Shibata-Koyama M., Isoda Y., Iida S., Masuda K., Satoh M., Kato K. Structural basis for improved efficacy of therapeutic antibodies on defucosylation of their Fc glycans. Genes Cells. 2011, 16:1071–1080.
Shields R.L., Lai J., Keck R., O’Connell L.Y., Hong K., Meng Y.G., Weikert S.H., Presta L.G. Lack of fucose on human IgG1 N-linked oligosaccharide improves binding to human Fcgamma RIII and antibody-dependent cellular toxicity. J. Biol. Chem. 2002, 277:26733–26740.
Jefferis R. Glycosylation as a strategy to improve antibody-based theraputics. Nat. Rev. Drug. Discov. 2009, 8:226–234.
Washburn N., Schwab I., Ortiz D., Bhatnagar N., Lansing J.C., Medeiros A., Tyler S., Mekala D., Cochran E., Sarvaiya H., Garofalo K., Meccariello R., Meador J.W., Rutitzky L., Schutles B.C., Ling L., Avery W., Nimmerjahn F., Manning A.M., Kaundinya G.V., Bosques C. Controlled tetra-Fc sialylation of IVIg results in a drug candidate with consistent enhanced anti-inflammatory activity. Proc. Natl. Acad. Sci. USA. 2015, 112: E1297–1306.
Wei Y., Li C., Huang W., Li B., Strome S., Wang L.-X. Glycoengineering of human IgG1-Fc through combined yeast expression and in vitro chemoenzymatic glycosylation. Biochemistry, 2008, 47:10294–10304.
Sjogren J., Struwe W.B., Cosgrave E.F.J., Rudd P.M., Stervander M., Allhorn M., Hollands A., Nizet V., Collin M. EndoS2 is a unique and conserved enzyme of sero-type M49 group A Streptococcus that hydrolyses N-linked glycans on IgG and α1-acid glycoprotein. Biochem. J. 2013, 455:107–118.
Lin C.-W., Tsai M.-H., Li S.-T., Tsai T.-I., Chu, K.-C., Liu Y.-C., Lai M.-Y., Wu C.-., Tseng Y.-C., Shivatare S.S., Wang C.-H., Chao P., Wang S.-Y., Shih H.-W., Zeng Y.-F., You T.-H., Liao J.-Y., Tu Y.-C., Lin Y.-S., Chuang H.-Y., Chen C.-L., Tsai C.-S., Huang C.-C., Lin N.-H., Ma C., Wu C.-Y., Wong C.-H. A common glycan structure on immunoglobulin G for enhancement of effector functions. Proc. Natl. Acad. Sci. USA, 2015, 112:10611–10616.
Parsons T.B., Struwe W.B., Gault J., Yamamoto K., Taylor T.A., Raj R., Wals K., Mohammed S., Robinson C.V., Benesch J.L.P., Davis B.G. Optimal synthetic glycosylation of a therapeutic antibody. Angew. Chem. Int. Ed. Engl. 2016, 55:2361–2367.
Li T., DiLillo D.J., Bournazos S., Giddens J.P., Ravetch J.V., Wang L.-X. Modulating IgG effector function by Fc glycan engineering. Proc. Natl. Acad. Sci. USA, 2017, 114:3485–3490.
Li C., Li T., Wang L.-X. Chemoenzymatic defucosylation of therapeutic antibodies for enhanced effector functions using bacterial α-fucosidases. Methods Mol. Biol. 2018, 1827: 367–380.
Kurogochi M., Mori M., Osumi K., Tojino M., Sugawara S., Takashima S., Hirose Y., Tsukimura W., Mizuno, M., Amano J., Matsuda A., Tomita M., Takayanagi A., Shoda S., Shirai T. Glycoengineered monoclonal antibodies with homogeneous glycan (M3, G0, G2, and A2) using a chemoenzymatic approach have different affinities for FcγRIIIa and variable antibody-dependent cellular cytotoxicity activities. Plos One. 2015, 10:e0132848.
Giddens J.P., Lomino J.V., DiLillo D.J., Ravetch J.V., Wang L.-X. Site-selective chemoenzymatic glycoengineering of Fab and Fc glycans of a therapeutic antibody. Proc. Natl. Acad. Sci. USA, 2018, 115:12023–12027.
Iwamoto M., Sekiguchi Y., Nakamura K., Kawaguchi Y., Honda T., Hasegawa J. Generation of efficient mutants of endoglycosidase from Streptococcus pyogenes and their application in a novel one-pot transglycosylation
reaction for antibody modification. Plos One. 2018, 13: e0193534.
Tang F., Yang Y., Tang Y., Tang S., Yang L., Sun B., Jiang B., Dong J., Liu H., Huang M., Geng M.-Y., Huang W. One-pot N-glycosylation remodeling of IgG with non-natural sialylglycopeptides enables glycosite-specific and dual-payload antibody-drug conjugates. Org. Biomol. Chem. 2016, 14:9501–9518.
Li T., Li C., Quan D.N., Bentley W.E., Wang L.-X. Site-specific immobilization of endoglycosidases for streamlined chemoenzymatic glycan remodeling of antibodies. Carbohydr. Res. 2018, 458–459:77–84.
Liu C.-P., Tsai T.-I., Cheng T., Shivatare V.S., Wu C.-Y., Wu C.-Y., Wong C.H. Glycoengineering of
antibody (Herceptin) through yeast expression and in vitro enzymatic glycosylation. Proc. Natl. Acad. Sci. USA. 2018, 115:720–725.
Eshima Y, Higuchi Y, Kinoshita T, Nakakita S, Takegawa K. Transglycosylation activity of glycosynthase mutants of endo-β-N-acetylglucosaminidase from Coprinopsis cinerea. Plos One. 2015, 10: e0132859.
Katoh T., Katayama T., Tomabechi Y., Nishikawa Y., Kumada J., Matsuzaki Y., Yamamoto K. Generation of a mutant Mucor hiemalis endoglycosidase that acts on core-fucosylated N-glycans. J. Biol. Chem, 2016, 291:23305–23317.
Shivatare S.S., Huang L.-Y., Zeng Y.-F., Liao J.-Y., You T.-H., Wang S.-Y., Cheng T., Chiu C.-W., Chao P., Chen L.-T., Tsai T.-I., Huang C.-C., Wu C.-Y., Lin N.-H., Wong C.-H. Development of glycosynthases with broad glycan specificity for the efficient glyco-remodeling of antibodies. Chem. Commun. 2018, 54:6161–6164.