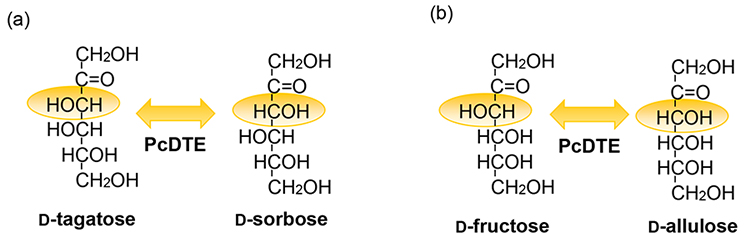
Fig. 1. P. cichorii D-TEの酵素反応 P. cichorii D-TE は(a)の異性化反応だけではなく、(b)の異性化反応も行うことができる。
D-fructose から D-allulose を生産することが可能。
2. 希少糖D-アルロースの生産
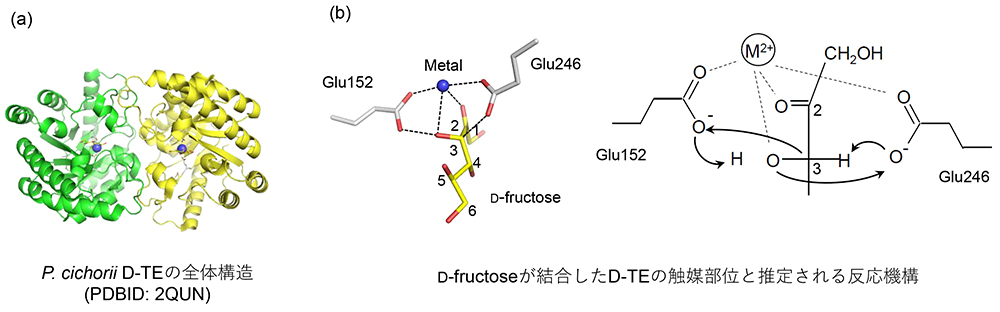
P. cichorii ST-24由来のD-TEがすべてのケトペントースやケトヘキソースに作用できることが明らかになり、大量生産が可能になった希少糖がD-アルロースである。D-アルロースは自然界に多量に存在するD-フルクトース(果糖)のC3エピマーであり、D-フルクトースにD-TEを作用させることで、D-フルクトースからD-アルロースへ約25%転換できることが明らかとなり、大量のD-アルロースの生産が可能となった。反応後平衡に達したD-フルクトース:D-アルロース=75:25の反応液からのD-アルロースの分離精製には、当初パン酵母を用いた資化反応によってD-フルクトースを除去する方法が取られてきたが、2000年頃に擬似移動式クロマト分離装置を導入することによって、純粋なD-アルロースの生産量を向上することができた4。また、D-アルロースを大量に作ることが出来るようになると、生産したD-アルロースを基質に用いて、新しい希少糖の生産へと展開していった。
Khan, A. R., Takahata, S., Okaya, H., Tsumura, T. and Izumori, K.; “D-Sorbose fermentation” from galactiol by Pseudomonas sp. ST 24. J. Ferment. Bioeng., 74(3), 149-152 (1992).
Izumori, K., Rahman, A. K., Okaya, H. and Tsumura, T.; A new enzyme, D-ketohexose 3-epimerase, from Pseudomonas sp. ST-24. Biosci. Biotech. Biochem., 57(6), 1037-1039 (1993).
Takeshita, K., Suga, A., Takada, G. and Izumori, K.; Mass production of D-psicose from D-fructose by a continuous bioreactor system using immobilized D-tagatose 3-epimerase. J. Biosci. Bioeng., 90(4), 453-455 (2000).
Yoshida, H., Yamada, M., Nishitani, T., Takada, G., Izumori, K. and Kamitori, S.; Crystal structures of D-tagatose 3-epimerase from Pseudomonas cichorii and its complexes with D-tagatose and D-fructose. J. Mol. Biol., 374(2), 443-453 (2007).
Kim, H. J., Hyun, E. K., Kim, Y. S., Lee, Y. J. and Oh, D. K.; Characterization of an Agrobacterium tumefaciens D-psicose 3-epimerase that converts D-fructose to D-psicose. Appl. Environ. Microbiol., 72(2), 981-985 (2006).
Jia, M., Mu, W., Chu, F., Zhang, X., Jiang, B., Zhou, L. L. and Zhang, T.; A D-psicose 3-epimerase with neutral pH optimum from Clostridium bolteae for D-psicose production: cloning, expression, purification, and characterization. Appl. Microbiol. Biotech., 98, 717-725 (2014).
Mu, W., Chu, F., Xing, Q., Yu, S., Zhou, L. and Jiang, B.; Cloning, expression, and characterization of a D-psicose 3-epimerase from Clostridium cellulolyticum H10. J. Agric. Food Chem., 59(14), 7785-7792 (2011).
Zhang, W., Li, H., Zhang, T., Jiang, B., Zhou, L. and Mu, W.; Characterization of a D-psicose 3-epimerase from Dorea sp. CAG317 with an acidic pH optimum and a high specific activity. J. Mol. Catal. B Enzym., 120, 68-74 (2015).
Zhu, Y., Men, Y., Bai, W., Li, X., Zhang, L., Sun, Y. and Ma, Y.; Overexpression of D-psicose 3-epimerase from Ruminococcus sp. in Escherichia coli and its potential application in d-psicose production. Biotechnol. Lett., 34, 1901-1906 (2012).
Zhang, W., Zhang, T., Jiang, B. and Mu, W.; Biochemical characterization of a D‐psicose 3‐epimerase from Treponema primitia ZAS‐1 and its application on enzymatic production of D‐psicose. J. Sci. Food Agric., 96(1), 49-56 (2016).
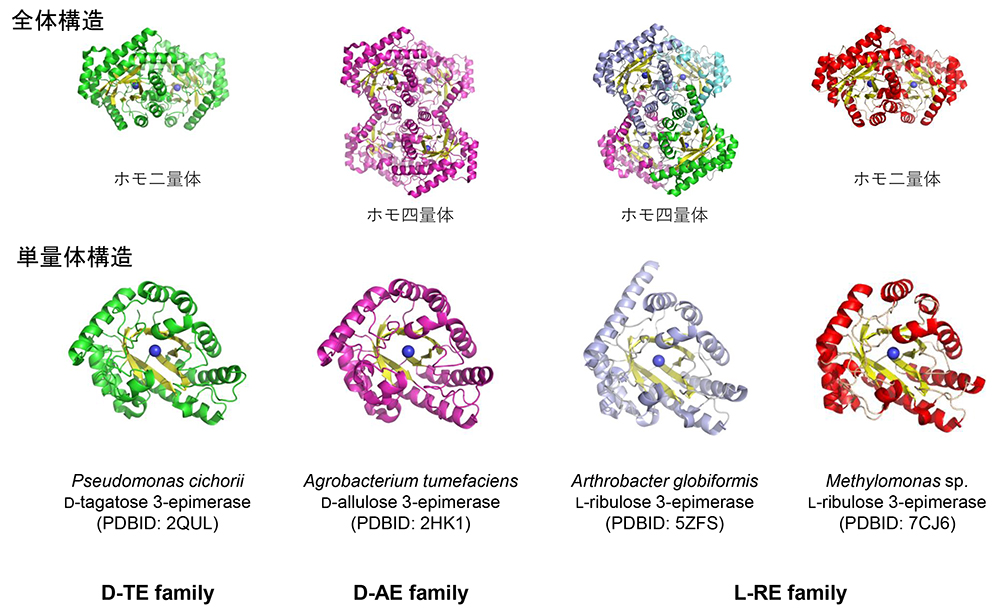
Yoshihara, A., Kozakai, T., Shintani, T., Matsutani, R., Ohtani, K., Iida, T., Akimitsu, K., Izumori, K. and Gullapalli, P. K.; Purification and characterization of D-allulose 3-epimerase derived from Arthrobacter globiformis M30, a GRAS microorganism. J. Biosci. Bioeng., 123(2), 170-176 (2017).
Yoshida, H., Yoshihara, A., Gullapalli, P. K., Ohtani, K., Akimitsu, K., Izumori, K. and Kamitori, S.; X-ray structure of Arthrobacter globiformis M30 ketose 3-epimerase for the production of D-allulose from D-fructose. Acta Crystallogr. F Struct. Biol. Commun., 74(10), 669-676 (2018).
Yoshida, H., Yoshihara, A., Kato, S., Mochizuki, S., Akimitsu, K., Izumori, K. and Kamitori, S.; Crystal structure of a novel homodimeric L‐ribulose 3‐epimerase from Methylomonus sp. FEBS Open Bio, 11(6), 1621-1637 (2021).
Chen, D., Chen, J., Liu, X., Guang, C., Zhang, W. and Mu, W.; Biochemical identification of a hyperthermostable L-ribulose 3-epimerase from Labedella endophytica and its application for D-allulose bioconversion. Int. J. Biol. Macromol., 189, 214-222 (2021).
Patel, S. N., Kaushal, G. and Singh, S. P.; A novel D-allulose 3-epimerase gene from the metagenome of a thermal aquatic habitat and D-allulose production by Bacillus subtilis whole-cell catalysis. Appl. Environ. Microbiol., 86(5), e02605-19 (2020).
Li, C., Li, L., Feng, Z., Guan, L., Lu, F. and Qin, H. M.; Two-step biosynthesis of D-allulose via a multienzyme cascade for the bioconversion of fruit juices. Food Chem., 357, 129746 (2021).
Laksmi, F. A., Nirwantono, R., Nuryana, I. and Agustriana, E.; Expression and characterization of thermostable D-allulose 3-epimerase from Arthrobacter psychrolactophilus (Ap DAEase) with potential catalytic activity for bioconversion of D-allulose from D-fructose. Int. J. Biol. Macromol., 214, 426-438 (2022).
Xie, X., Tian, Y., Ban, X., Li, C., Yang, H. and Li, Z.; Crystal structure of a novel homodimeric D-allulose 3-epimerase from a Clostridia bacterium. Acta Crystallogr. D Struct. Biol., 78(9), 1180-1191 (2022).
Qi, H., Wang, T., Li, H., Li, C., Guan, L., Liu, W., Wang, J., Lu, F., Mao, S. and Qin, H. M.; Sequence-and structure-based mining of thermostable D-allulose 3-epimerase and computer-guided protein engineering to improve enzyme activity. J. Agric. Food Chem., 71(47), 18431-18442 (2023).
Zhang, W., Zhang, Y., Huang, J., Chen, Z., Zhang, T., Guang, C. and Mu, W.; Thermostability improvement of the D-allulose 3-epimerase from Dorea sp. CAG317 by site-directed mutagenesis at the interface regions. J. Agric. Food Chem., 66(22), 5593-5601 (2018).
Zhao, J., Chen, J., Wang, H., Guo, Y., Li, K. and Liu, J.; Enhanced thermostability of D-psicose 3-epimerase from Clostridium bolteae through rational design and engineering of new disulfide bridges. Int. J. Mol. Sci., 22(18), 10007 (2021).
Zhu, Z., Li, L., Zhang, W., Li, C., Mao, S., Lu, F. and Qin, H. M.; Improving the enzyme property of D-allulose 3-epimerase from a thermophilic organism of Halanaerobium congolense through rational design. Enzyme Microb. Technol., 149, 109850 (2021).
Li, C., Gao, X., Li, H., Wang, T., Lu, F., and Qin, H. M.; Growth‐Coupled Evolutionary Pressure Improving Epimerases for D‐Allulose Biosynthesis Using a Biosensor‐Assisted In Vivo Selection Platform. Adv. Sci. (Weinh), 11(14), 2306478 (2024).
Kim, K., Kim, H. J., Oh, D. K., Cha, S. S. and Rhee, S.; Crystal structure of D-psicose 3-epimerase from Agrobacterium tumefaciens and its complex with true substrate D-fructose: a pivotal role of metal in catalysis, an active site for the non-phosphorylated substrate, and its conformational changes. J. Mol. Biol., 361(5), 920-931 (2006).
Chan, H. C., Zhu, Y., Hu, Y., Ko, T. P., Huang, C. H., Ren, F., Chen, C. C., Ma, Y., Guo, R.T. and Sun, Y.; Crystal structures of D-psicose 3-epimerase from Clostridium cellulolyticum H10 and its complex with ketohexose sugars. Protein Cell, 3, 123-131 (2012).
Yoshida, H., Yoshihara, A., Ishii, T., Izumori, K. and Kamitori, S.; X-ray structures of the Pseudomonas cichorii D-tagatose 3-epimerase mutant form C66S recognizing deoxy sugars as substrates. Appl. Microbiol. Biotechnol., 100(24), 10403-10415 (2016).
Xie, X., Tian, Y., Ban, X., Li, C., Yang, H. and Li, Z.; Crystal structure of a novel homodimeric D-allulose 3-epimerase from a Clostridia bacterium. Acta Crystallogr. D Struct. Biol., 78(9), 1180-1191 (2022).
Li, C., Gao, X., Qi, H., Zhang, W., Li, L., Wei, C., ... & Qin, H. M.; Substantial Improvement of an Epimerase for the Synthesis of D‐Allulose by Biosensor‐Based High‐Throughput Microdroplet Screening. Angew. Chem. Int. Ed. Engl., 135(10), e202216721 (2023).
Guo, D., Wang, Z., Wei, W., Song, W., Wu, J., Wen, J., Hu, G., Li, X., Gao, C., Chen, X. and Liu, L.; Rational design improves both thermostability and activity of a new D-tagatose 3-epimerase from Kroppenstedtia eburnean to produce D-allulose. Enzyme Microb. Technol., 178, 110448 (2024).
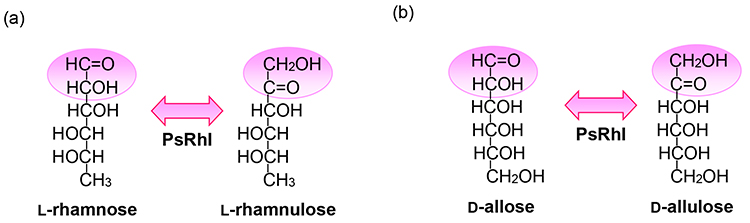
Bhuiyan, S. H., Itami, Y. and Izumori, K.; Isolation of an L-rhamnose isomerase-constitutive mutant of Pseudomonas sp. strain LL172: purification and characterization of the enzyme. J. Ferment. Bioeng., 84(4), 319-323 (1997).
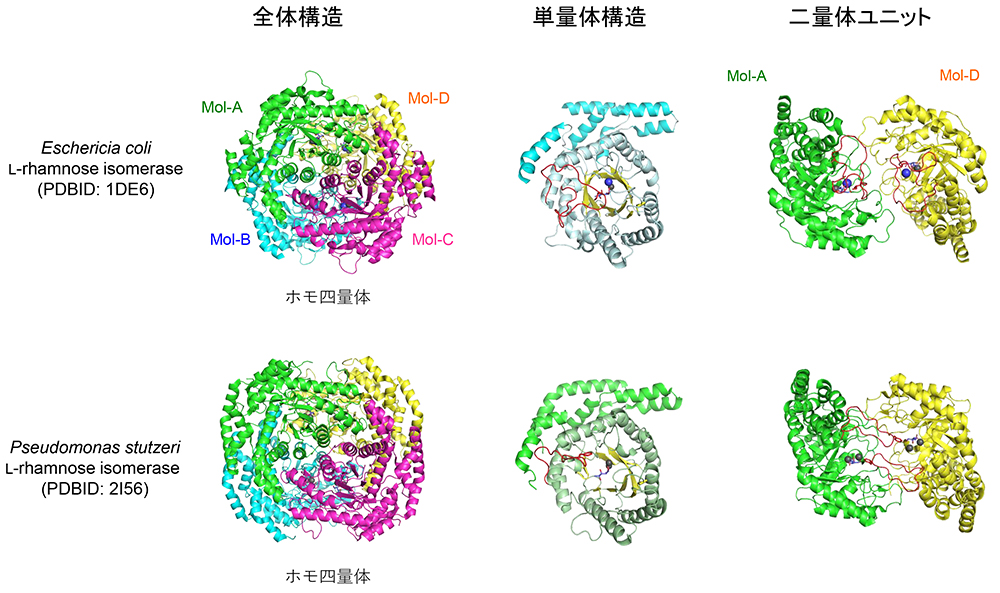
Korndörfer, I. P., Fessner, W. D. and Matthews, B. W.; The structure of rhamnose isomerase from Escherichia coli and its relation with xylose isomerase illustrates a change between inter and intra-subunit complementation during evolution. J. Mol. Biol., 300(4), 917-933 (2000).
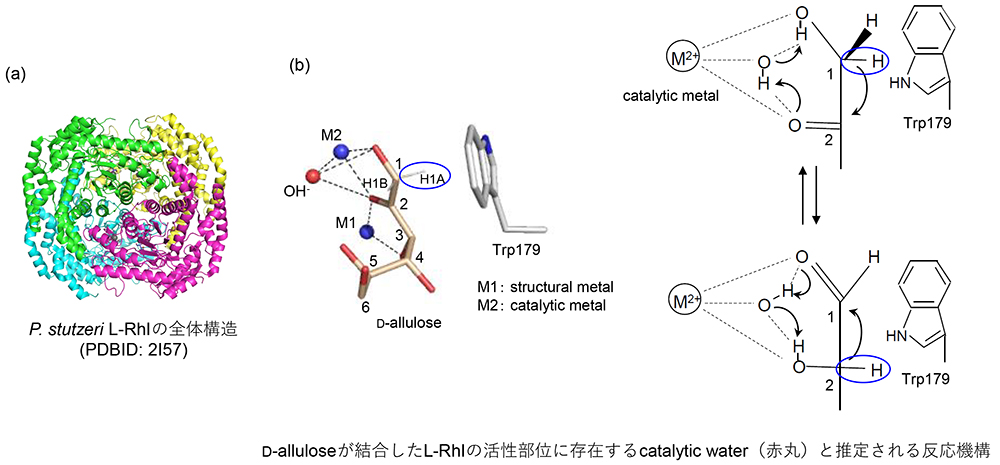
Yoshida, H., Yamada, M., Ohyama, Y., Takada, G., Izumori, K. and Kamitori, S.; The structures of L-rhamnose isomerase from Pseudomonas stutzeri in complexes with L-rhamnose and D-allose provide insights into broad substrate specificity. J. Mol. Biol., 365(5), 1505-1516 (2007).
Leang, K., Takada, G., Fukai, Y., Morimoto, K., Granström, T. B. and Izumori, K.; Novel reactions of L-rhamnose isomerase from Pseudomonas stutzeri and its relation with D-xylose isomerase via substrate specificity. Biochim. Biophys. Acta Gen. Subj., 1674(1), 68-77 (2004).
Prabhu, P., Doan, T. N. T., Tiwari, M., Singh, R., Kim, S. C., Hong, M. K., Kang, Y. C., Kang, L. W. and Lee, J. K.; Structure‐based studies on the metal binding of two‐metal‐dependent sugar isomerases. FEBS J., 281(15), 3446-3459 (2014).
Yoshida, H., Yamamoto, N., Kurahara, L. H., Izumori, K. and Yoshihara, A.; X-ray structure and characterization of a probiotic Lactobacillus rhamnosus Probio-M9 L-rhamnose isomerase. Appl. Microbiol. Biotechnol., 108(1), 249 (2024).
Yoshida, H., Izumori, K. and Yoshihara, A.; L-rhamnose isomerase: a crucial enzyme for rhamnose catabolism and conversion of rare sugars. Appl. Microbiol. Biotechnol., 108(1), 1-15 (2024).