Ohtsubo K, Marth JD. Glycosylation in cellular mechanisms of health and disease. Cell. 2006;126(5):855–867.
Paulson JC, Blixt O, Collins BE. Sweet spots in functional glycomics. Nat. Chem. Biol. 2006;2(5):238–248.
Pinho SS, Reis CA. Glycosylation in cancer: Mechanisms and clinical implications. Nat. Rev. Cancer. 2025;15(9):540–555.
Varki A. Biological roles of glycans. Glycobiology. 2017;27(1):3–49.
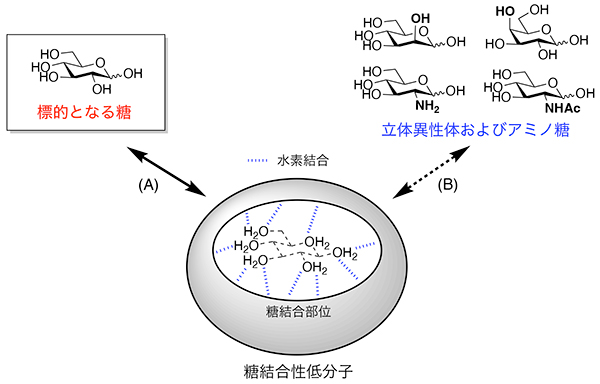
Davis AP. Synthetic lectins. Org. Biomol. Chem. 2009;7(18):3629–3638.
Oki T, Konishi M, Tomatsu K, Tomita K, Saitoh K, Tsunakawa M, Nishio M, Miyaki T, Kawaguchi H. Pradimicin, a novel class of potent antifungal antibiotics. J. Antibiot. 1988;41(11):1701–1704.
Fukagawa Y, Ueki T, Numata K, Oki T. Pradimicins and benanomicins, sugar-recognizing antibiotics: Their novel mode of antifungal action and conceptual significance. Actinomycetol. 1993;7(1):1–22.
Fujikawa K, Tsukamoto Y, Oki T, Lee YC. Spectroscopic studies on the interaction of pradimicin BMY-28864 with mannose derivatives. Glycobiology. 1998;8(4):407–414.
Ueki T, Numata K, Sawada Y, Nakajima T, Fukagawa Y, Oki T. Studies on the mode of antifungal action of pradimicin antibiotics. I. Lectin-mimic binding of BMY-28864 to yeast mannan in the presence of calcium. J. Antibiot. 1993;46(1):149–161.
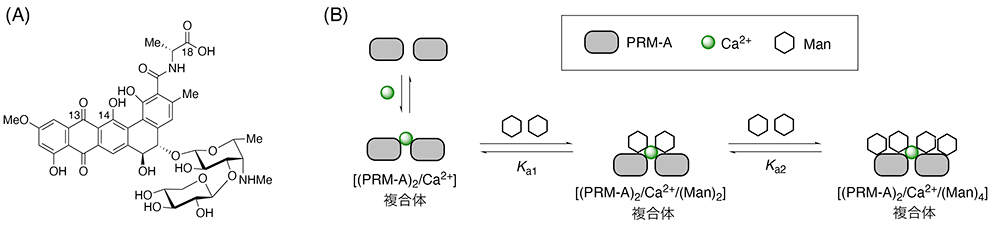
Nakagawa Y, Masuda Y, Yamada K, Doi T, Takegoshi K, Igarashi Y, Ito Y. Solid-state NMR spectroscopic analysis of the Ca2+-dependent mannose binding of pradimicin A. Angew. Chem. Int. Ed. Engl. 2011;50(27):6084–6088.
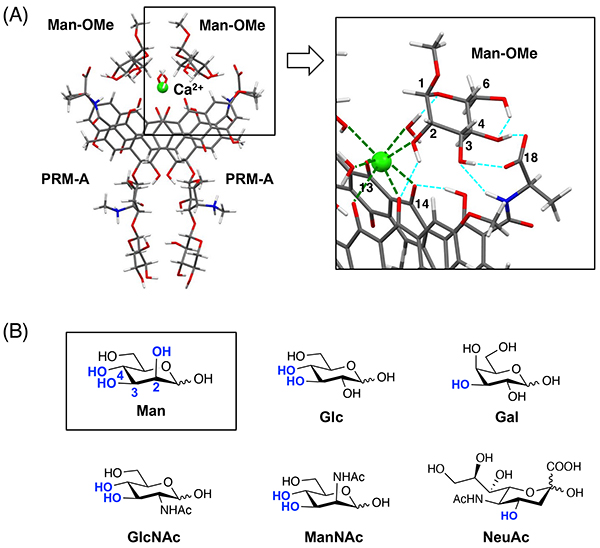
Nakagawa Y, Doi T, Masuda Y, Takegoshi K, Igarashi Y, Ito Y. Mapping of the primary mannose binding site of pradimicin A. J. Am. Chem. Soc. 2011;133(43):17485–17493.
Nakagawa Y, Doi T, Taketani T, Takegoshi K, Igarashi Y, Ito Y. Mannose-binding geometry of pradimicin A. Chem. Eur. J. 2013;19(32):10516–10525.
Takegoshi K, Nakamura S, Terao T. 13C–1H dipolar-assisted rotational resonance in magic-angle spinning NMR. Chem. Phys. Lett. 2001;344(5–6):631–637.
Takegoshi K, Nakamura S, Terao T. 13C–1H dipolar-driven 13C–13C recoupling without 13C rf irradiation in nuclear magnetic resonance of rotating solids. J. Chem. Phys. 2003;118(5):2325–2341.
Nakagawa Y, Doi T, Takegoshi K, Sugahara T, Akase D, Aida M, Tsuzuki K, Watanabe Y, Tomura T, Ojika M, Igarashi Y, Hashizume D, Ito Y. Molecular basis of mannose recognition by pradimicins and their application to microbial cell surface imaging. Cell Chem. Biol. 2019;26(7):950–959.
Ueki T, Oka M, Fukagawa Y, Oki T. Studies on the mode of antifungal action of pradimicin antibiotics. III. Spectrophotometric sequence analysis of the ternary complex formation of BMY-28864 with D-mannopyranoside and calcium. J. Antibiot. 1993;46(3):465–477.
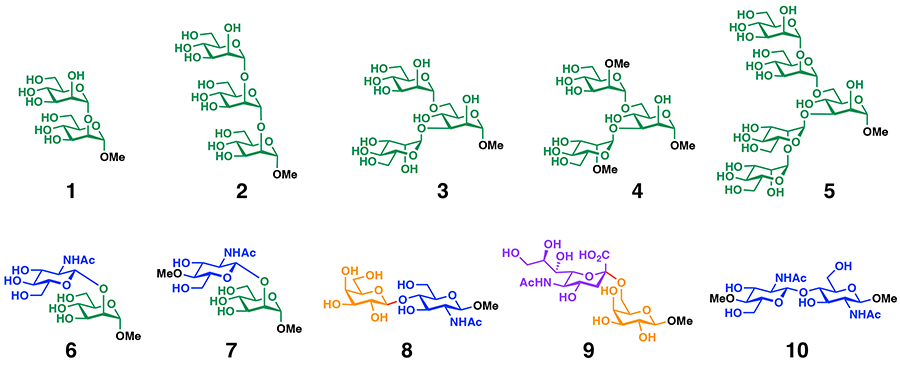
Nakagawa Y, Yamaji F, Miyanishi W, Ojika M, Igarashi Y, Ito Y. Binding evaluation of pradimicins for oligomannose motifs from fungal mannans. Bull. Chem. Soc. Jpn. 2021;94(3):732–754.
Nakagawa Y, Fujii M, Ito N, Ojika M, Akase D, Aida M, Kinoshita T, Sakurai Y, Yasuda J, Igarashi Y, Ito Y. Molecular basis of N-glycan recognition by pradimicin A and its potential as a SARS-CoV-2 entry inhibitor. Bioorg. Med. Chem. 2024;105:117732.
Nakagawa Y, Kakihara S, Tsuzuki K, Ojika M, Igarashi Y, Ito Y. A pradimicin-based staining dye for glycoprotein detection. J. Nat. Prod. 2021;84(9):2496–2501.
Miyanishi W, Ojika M, Akase D, Aida M, Igarashi Y, Ito Y, Nakagawa Y. D-mannose binding aggregation property, and antifungal activity of amide derivatives of pradimicin A. Bioorg. Med. Chem. 2022;55:116590.
Crispin M, Ward AB, Wilson IA. Structure and immune recognition of the HIV glycan shield. Annu. Rev. Biophys. 2018;47:499–523.
Watanabe Y, Bowden TA, Wilson IA, Crispin M. Exploitation of glycosylation in enveloped virus pathobiology. Biochim. Biophys. Acta. Gen. Subj. 2019;1863(10):1480–1497.
François KO, Balzarini J. Potential of carbohydrate-binding agents as therapeutics against enveloped viruses. Med. Res. Rev. 2012;32(2):349–387.
Piotrowski JS, Okada H, Lu F, Li SC, Hinchman L, Ranjan A, Smith DL, Higbee AJ, Ulbrich A, Coon JJ, Deshpande R, Bukhman YV, Mcllwain S, Ong IM, Myers CL, Boone C, Landick R, Ralph J, Kabbage M, Ohya Y. Plant-derived antifungal agent poacic acid targets β-1,3-glucan. Proc. Natl. Acad. Sci. USA. 2015;112(12):E1490–E1497.
Yona A, Fridman M. Poacic acid, a plant-derived stilbenoid, augments cell wall chitin production, but its antifungal activity is hindered by this polysaccharide and by fungal essential metals. Biochemistry. 2024;63(8):1051–1065.
Tromans RA, Carter TS, Chabanne L, Crump MP, Li H, Matlock JV, Orchard MG, Davis AP. A biomimetic receptor for glucose. Nat. Chem. 2019;11(1):52–56.
Lozano V, Aguado L, Hoorelbeke B, Renders M, Camarasa MJ, Schols D, Balzarini J, San-Félix A, Pérez-Pérez MJ. Targeting HIV entry through interaction with envelope glycoprotein 120 (gp120): Synthesis and antiviral evaluation of 1,3,5-triazines with aromatic amino acids. J. Med. Chem. 2011;54(15):5335–5348.
Rivero-Buceta E, Carrero P, Casanova E, Doyagüez EG, Madrona A, Quesada E, Peréz-Pérez MJ, Mateos R, Bravo L, Mathys L, Noppen S, Kiselev E, Marchand C, Pommier Y, Liekens S, Balzarini J, Camarasa MJ, San-Félix A. Anti-HIV-1 activity of a tripodal receptor that recognizes mannose-oligomers. Eur. J. Med. Chem. 2015;106:132–143.
Francesconi O, Nativi C, Gabrielli G, De Simone I, Noppen S, Balzarini J, Liekens S, Roelens S. Antiviral activity of synthetic aminopyrrolic carbohydrate binding agents: targeting the glycans of viral gp120 to inhibit HIV entry. Chem. Eur. J. 2015;21(28):10089–10093.