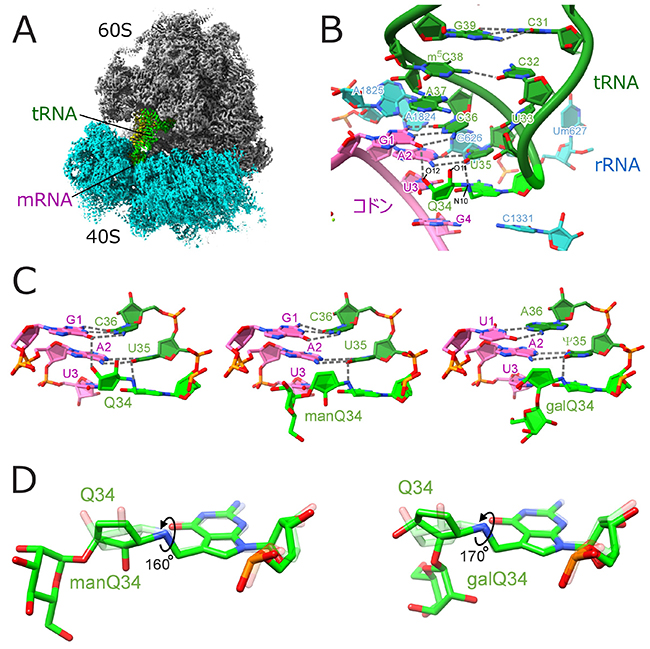
図 3. 糖付加Q修飾はコドン-アンチコドン対合の主溝に結合する
A.クライオ電子顕微鏡によるヒト80Sリボソーム複合体の構造
B. Aサイトにおけるコドンアンチコドン対合を拡大
C. GAUコドンを認識するQ修飾(左)とmanQ修飾(中)、UAUコドンを認識するgalQ修飾(右)の構造。Qのシクロペンテン環はコドンアンチコドン対合に主溝側から結合し、複数の水素結合を形成するが、糖付加Q修飾は水素結合による認識が見られない。
D. マンノース(左)およびガラクトース(右)の付加はQのシクロペンテン環をそれぞれ160°と170°回転させ、主溝結合能を低下させることで、翻訳速度を制御する。
Cappannini, A., Ray, A., Purta, E., Mukherjee, S., Boccaletto, P., Moafinejad, S.N., Lechner, A., Barchet, C., Klaholz, B.P., Stefaniak, F., and Bujnicki, J.M. (2024). MODOMICS: a database of RNA modifications and related information. 2023 update. Nucleic Acids Res 52, D239-D244.
Suzuki, T. (2021). The expanding world of tRNA modifications and their disease relevance. Nat Rev Mol Cell Biol 22, 375-392.
Miyauchi, K., Kimura, S., Akiyama, N., Inoue, K., Ishiguro, K., Vu, T., Srisuknimit, V., Koyama, K., Hayashi, G., Soma, A., et al. (2024). A tRNA modification with aminovaleramide facilitates AUA decoding in protein synthesis. Nat Chem Biol, in press.
Ohira, T., Minowa, K., Sugiyama, K., Yamashita, S., Sakaguchi, Y., Miyauchi, K., Noguchi, R., Kaneko, A., Orita, I., Fukui, T., et al. (2022). Reversible RNA phosphorylation stabilizes tRNA for cellular thermotolerance. Nature 605, 372-379.
Chujo, T., and Tomizawa, K. (2021). Human transfer RNA modopathies: diseases caused by aberrations in transfer RNA modifications. FEBS J 288, 7096-7122.
Asano, K., Suzuki, T., Saito, A., Wei, F.Y., Ikeuchi, Y., Numata, T., Tanaka, R., Yamane, Y., Yamamoto, T., Goto, T., et al. (2018). Metabolic and chemical regulation of tRNA modification associated with taurine deficiency and human disease. Nucleic Acids Res 46, 1565-1583.
Suzuki, T., Nagao, A., and Suzuki, T. (2011). Human mitochondrial tRNAs: biogenesis, function, structural aspects, and diseases. Annual review of genetics 45, 299-329.
de Crecy-Lagard, V., Hutinet, G., Cediel-Becerra, J.D.D., Yuan, Y., Zallot, R., Chevrette, M.G., Ratnayake, R., Jaroch, M., Quaiyum, S., and Bruner, S. (2024). Biosynthesis and function of 7-deazaguanine derivatives in bacteria and phages. Microbiol Mol Biol Rev 88, e0019923.
Kasai, H., Nakanishi, K., Macfarlane, R.D., Torgerson, D.F., Ohashi, Z., McCloskey, J.A., Gross, H.J., and Nishimura, S. (1976). Letter: The structure of Q* nucleoside isolated from rabbit liver transfer ribonucleic acid. J Am Chem Soc 98, 5044-5046.
Lairson, L.L., Henrissat, B., Davies, G.J., and Withers, S.G. (2008). Glycosyltransferases: structures, functions, and mechanisms. Annu Rev Biochem 77, 521-555.
Moremen, K.W., Ramiah, A., Stuart, M., Steel, J., Meng, L., Forouhar, F., Moniz, H.A., Gahlay, G., Gao, Z., Chapla, D., et al. (2018). Expression system for structural and functional studies of human glycosylation enzymes. Nat Chem Biol 14, 156-162.
Zhao, X., Ma, D., Ishiguro, K., Saito, H., Akichika, S., Matsuzawa, I., Mito, M., Irie, T., Ishibashi, K., Wakabayashi, K., et al. (2023). Glycosylated queuosines in tRNAs optimize translational rate and post-embryonic growth. Cell 186, 5517-5535 e5524.
Panneerselvam, K., and Freeze, H.H. (1996). Mannose enters mammalian cells using a specific transporter that is insensitive to glucose. J Biol Chem 271, 9417-9421.
Milo, R., Jorgensen, P., Moran, U., Weber, G., and Springer, M. (2010). BioNumbers--the database of key numbers in molecular and cell biology. Nucleic Acids Res 38, D750-753.
Kirkman, H.N., Jr. (1992). Erythrocytic uridine diphosphate galactose in galactosaemia. J Inherit Metab Dis 15, 4-16.
Keevill, N.J., Holton, J.B., and Allen, J.T. (1994). UDP-glucose and UDP-galactose concentrations in cultured skin fibroblasts of patients with classical galactosaemia. J Inherit Metab Dis 17, 23-26.
Nedialkova, D.D., and Leidel, S.A. (2015). Optimization of Codon Translation Rates via tRNA Modifications Maintains Proteome Integrity. Cell 161, 1606-1618.
Fergus, C., Barnes, D., Alqasem, M.A., and Kelly, V.P. (2015). The queuine micronutrient: charting a course from microbe to man. Nutrients 7, 2897-2929.
Gunduz, U., Elliott, M.S., Seubert, P.H., Houghton, J.A., Houghton, P.J., Trewyn, R.W., and Katze, J.R. (1992). Absence of tRNA-guanine transglycosylase in a human colon adenocarcinoma cell line. Biochim Biophys Acta 1139, 229-238.
Baranowski, W., Dirheimer, G., Jakowicki, J.A., and Keith, G. (1994). Deficiency of queuine, a highly modified purine base, in transfer RNAs from primary and metastatic ovarian malignant tumors in women. Cancer Res 54, 4468-4471.
Huang, B.S., Wu, R.T., and Chien, K.Y. (1992). Relationship of the queuine content of transfer ribonucleic acids to histopathological grading and survival in human lung cancer. Cancer Res 52, 4696-4700.
Emmerich, B., Zubrod, E., Weber, H., Maubach, P.A., Kersten, H., and Kersten, W. (1985). Relationship of queuine-lacking transfer RNA to the grade of malignancy in human leukemias and lymphomas. Cancer Res 45, 4308-4314.
Zhang, J., Lu, R., Zhang, Y., Matuszek, Z., Zhang, W., Xia, Y., Pan, T., and Sun, J. (2020). tRNA Queuosine Modification Enzyme Modulates the Growth and Microbiome Recruitment to Breast Tumors. Cancers (Basel) 12, 628.
Blum, A., Wang, P., and Zenklusen, J.C. (2018). SnapShot: TCGA-Analyzed Tumors. Cell 173, 530.
Errichiello, E., Lecca, M., Vantaggiato, C., Motta, Z., Zanotta, N., Zucca, C., Bertuzzo, S., Piubelli, L., Pollegioni, L., and Bonaglia, M.C. (2024). Further evidence supporting the role of GTDC1 in glycine metabolism and neurodevelopmental disorders. Eur J Hum Genet 32, 920-927.
Aksoy, I., Utami, K.H., Winata, C.L., Hillmer, A.M., Rouam, S.L., Briault, S., Davila, S., Stanton, L.W., and Cacheux, V. (2017). Personalized genome sequencing coupled with iPSC technology identifies GTDC1 as a gene involved in neurodevelopmental disorders. Hum Mol Genet 26, 367-382.
Flynn, R.A., Pedram, K., Malaker, S.A., Batista, P.J., Smith, B.A.H., Johnson, A.G., George, B.M., Majzoub, K., Villalta, P.W., Carette, J.E., and Bertozzi, C.R. (2021). Small RNAs are modified with N-glycans and displayed on the surface of living cells. Cell 184, 3109-3124.e22.