Cowman MK, Lee H-G, Schwertfeger KL, McCarthy JB, Turley EA. The content and size of Hyaluronan in biological fluids and tissues. Front Immunol. 2015:6:261. https://doi.org/10.3389/fimmu.2015.00261.
Itano N. Simple primary structure, complex turnover regulation and multiple roles of hyaluronan. J Biochem. 2008:144(2):131–137. https://doi.org/10.1093/jb/mvn046.
D’Agostino A, Stellavato A, Corsuto L, Diana P, Filosa R, La Gatta A, De Rosa M, Schiraldi C. Is molecular size a discriminating factor in hyaluronan interaction with human cells? Carbohydr Polym. 2017:157:21–30. https://doi.org/10.1016/j.carbpo l.2016.07.125.
Weigel PH, Baggenstoss BA. What is special about 200 kDa hyaluronan that activates hyaluronan receptor signaling? Glycobiology. 2017:27(9):868–877. https://doi.org/10.1093/glycob/cwx039.
Zakusilo FT, Kerry, O’Banion M, Gelbard HA, Seluanov A, Gorbunova V. Matters of size: roles of hyaluronan in CNS aging and dis- ease. Ageing Res Rev. 2021:72:101485. https://doi.org/10.1016/j.arr.2021.101485.
Yamaguchi Y. Lecticans: organizers of the brain extracellular matrix. Cell Mol Life Sci. 2000:57(2):276–289. https://doi.org/10.1007/PL00000690.
Miyata S, Kitagawa H. Formation and remodeling of the brain extra- cellular matrix in neural plasticity: roles of chondroitin sulfate and hyaluronan. Biochim Biophys Acta Gen Subj. 2017:1861(10): 2420–2434. https://doi.org/10.1016/j.bbagen.2017.06.010.
Deepa SS, Carulli D, Galtrey C, Rhodes K, Fukuda J, Mikami T, Sugahara K, Fawcett JW. Composition of perineuronal net extracellular matrix in rat brain: a different disaccharide composition for the net-associated proteoglycans. J Biol Chem. 2006:281(26): 17789–17800. https://doi.org/10.1074/jbc.M600544200.
Takechi M, Oshima K, Nadano D, Kitagawa H, Matsuda T, Miyata S. A pericellular hyaluronan matrix is FD for the morpho- logical maturation of cortical neurons. Biochim Biophys Acta Gen Subj. 2020:1864(10):129679. https://doi.org/10.1016/j.bbagen.2020.129679.
Pizzorusso T, Medini P, Berardi N, Chierzi S, Fawcett JW, Maffei L. Reactivation of ocular dominance plasticity in the adult visual cortex. Science. 2002:298(5596):1248–1251. https://doi.org/10.1126/science.1072699.
Zimmermann DR, Dours-Zimmermann MT. Extracellular matrix of the central nervous system: from neglect to challenge. Histochem Cell Biol. 2008:130(4):635–653. https://doi.org/10.1007/s00418-008-0485-9.
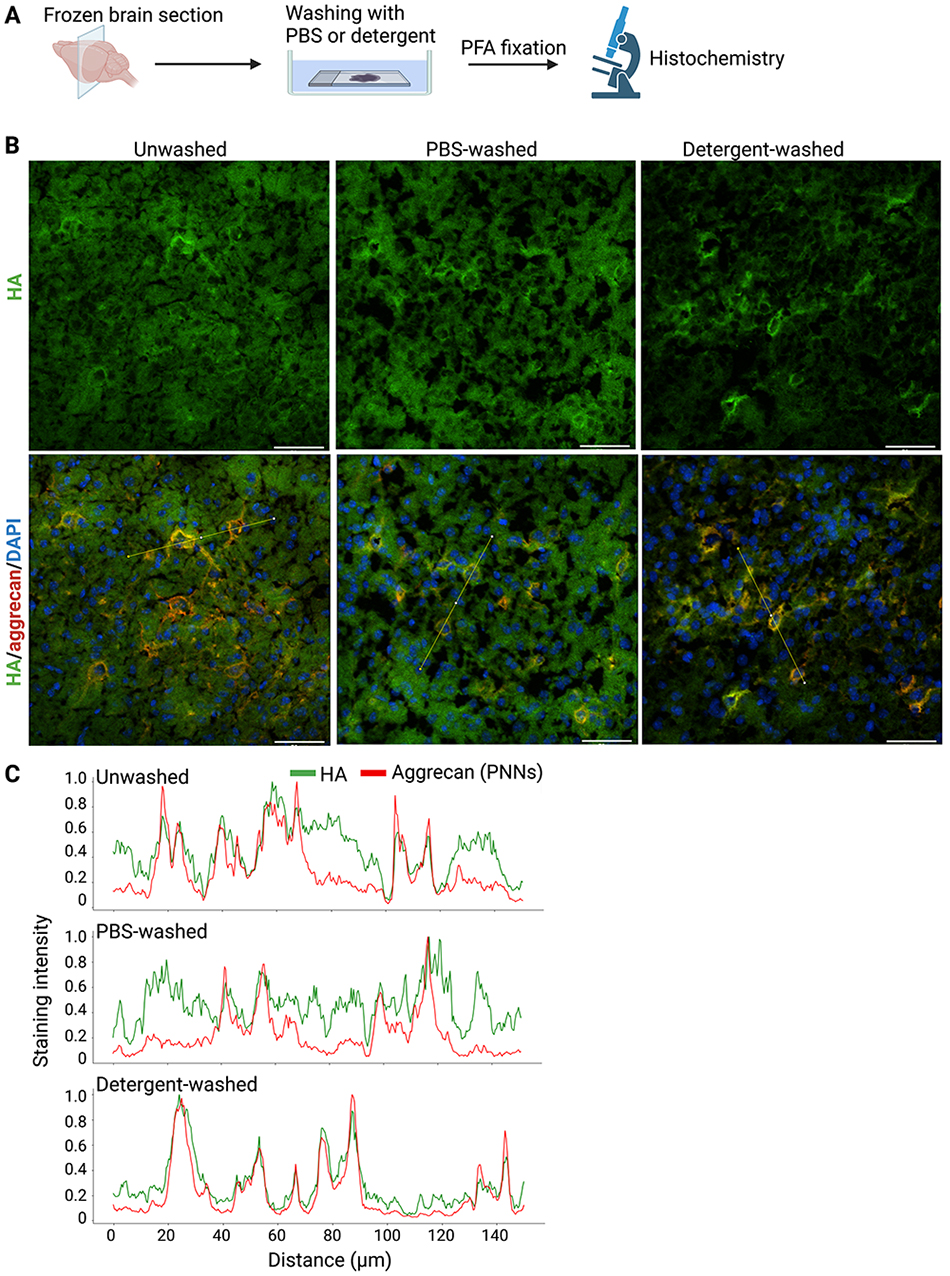
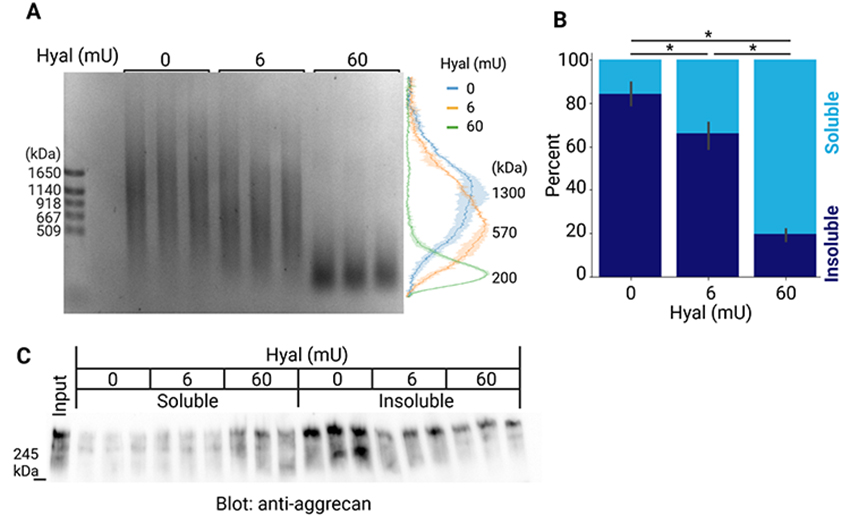
Sugitani K, Egorova D, Mizumoto S, Nishio S, Yamada S, Kitagawa H, Oshima K, Nadano D, Matsuda T, Miyata S. Hyaluronan degradation and release of a hyaluronan-aggrecan complex from perineuronal nets in the aged mouse brain. Biochim Biophys Acta Gen Subj. 2021:1865(2):129804. https://doi.org/10.1016/j.bbagen.2020.129804.
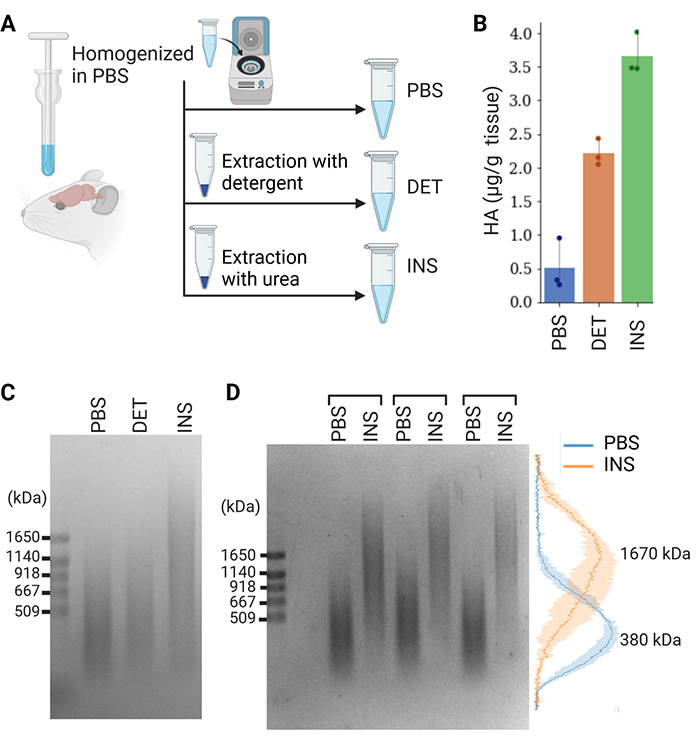
Egorova D, Nomura Y, Miyata S. Impact of hyaluronan size on localization and solubility of the extracellular matrix in the mouse brain. Glycobiology. 2023:33(8):615-625. https://doi.org/10.1093/glycob/cwad022.
Carulli D, Pizzorusso T, Kwok JCF, Putignano E, Poli A, Forostyak S, Andrews MR, Deepa SS, Glant TT, Fawcett JW. Animals lacking link protein have attenuated perineuronal nets and persistent plasticity. Brain. 2010:133(8):2331–2347. https://doi.org/10.1093/brain/awq145.
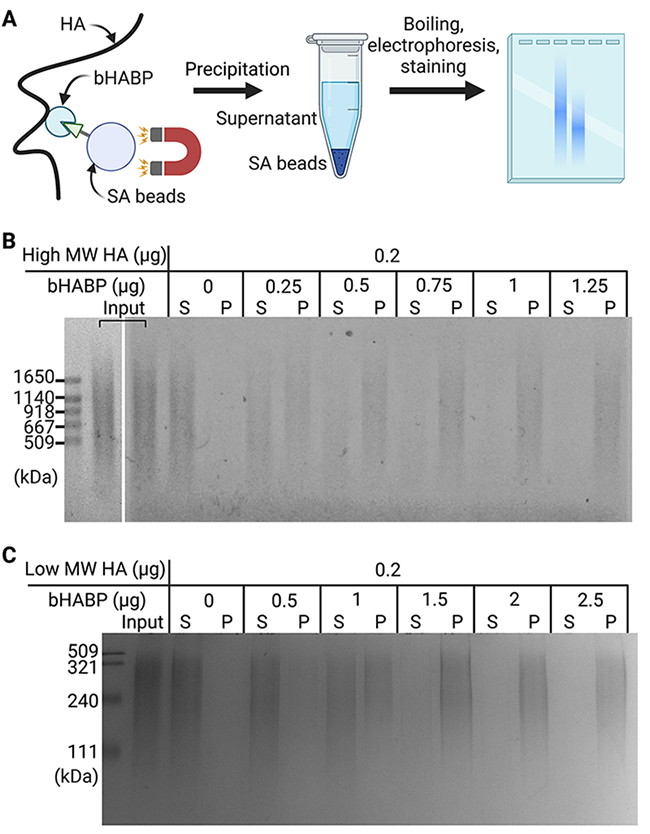
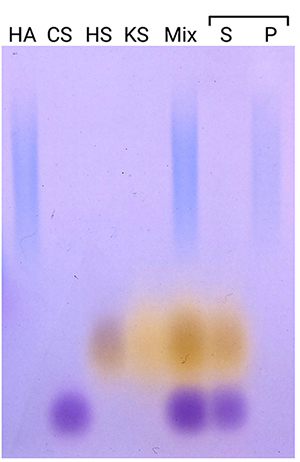
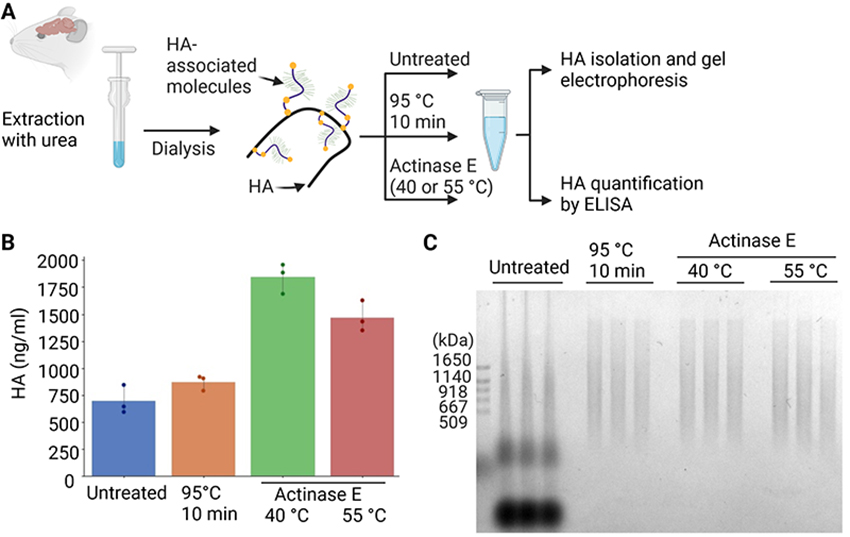
Rivas F, Erxleben D, Smith I, Rahbar E, DeAngelis PL, Cowman MK, Hall AR. Methods for isolating and analyzing physiological hyaluronan: a review. Am J Physiol Cell Physiol. 2022:322(4):C674–C687. https://doi.org/10.1152/ajpcell.00019.2022.
Fawcett JW, Fyhn M, Jendelova P, Kwok JCF, Ruzicka J, Sorg BA. The extracellular matrix and perineuronal nets in memory. Mol Psychiatry. 2022:27(8):3192–3203. https://doi.org/10.1038/s41380-022-01634-3.
Gogolla N, Caroni P, Lüthi A, Herry C. Perineuronal nets protect fear memories from erasure. Science. 2009:325(5945):1258–1261. https://doi.org/10.1126/science.1174146.
Thompson EH, Lensjø KK, Wigestrand MB, Malthe-Sørenssen A, Hafting T, Fyhn M. Removal of perineuronal nets disrupts recall of a remote fear memory. Proc Natl Acad Sci U S A. 2018:115(3): 607–612. https://doi.org/10.1073/pnas.1713530115.
Tsien RY. Very long-term memories may be stored in the pattern of holes in the perineuronal net. Proc Natl Acad Sci U S A. 2013:110(30): 12456–12461. https://doi.org/10.1073/pnas.1310158110.
de Vivo L, Landi S, Panniello M, Baroncelli L, Chierzi S, Mariotti L, Spolidoro M, Pizzorusso T, Maffei L, Ratto GM. Extracellular matrix inhibits structural and functional plasticity of dendritic spines in the adult visual cortex. Nat Commun. 2013:4(1):1484. https://doi.org/10.1038/ncomms2491.
Sorg BA, Berretta S, Blacktop JM, Fawcett JW, Kitagawa H, Kwok JCF, Miquel M. Casting a wide net: role of perineuronal nets in neural plasticity. J Neurosci. 2016:36(45):11459–11468. https://doi.org/10.1523/JNEUROSCI.2351-16.2016.