Lindahl, U., Bäckström, G., Thunberg, L., and Leder, I. G. (1980) Evidence for a 3-O-sulfated D-glucosamine residue in the antithrombin-binding sequence of heparin. Proc. Natl. Acad. Sci. USA77, 6551-6555
Petitou, M., Casu, B., and Lindahl, U. (2003) 1976-1983, a critical period in the history of heparin: the discovery of the antithrombin binding site. Biochimie85, 83-89
Petitou, M., Duchaussoy, P., Lederman, I., Choay, J., and Sinaÿ, P. (1988) Binding of heparin to antithrombin III: a chemical proof of the critical role played by a 3-sulfated 2-amino-2-deoxy-D-glucose residue. Carbohydr. Res. 179, 163-172
Liu, J., Shworak, N. W., Fritze, L. M. S., Edelberg, J. M., and Rosenberg, R. D. (1996) Purification of heparan sulfate D-glucosaminyl 3-O-sulfotransferase. J. Biol. Chem. 271, 27072-27082
Shworak, N. W., Liu, J., Fritze, L. M. S., Schwartz, J. J., Zhang, L., Logeart, D., and Rosenberg, R. D. (1997) Molecular cloning and expression of mouse and human cDNAs encoding heparan sulfate D-glucosaminyl 3-O-sulfotransferase. J. Biol. Chem. 272, 28008-28019
Shworak, N. W., Liu, J., Petros, L. M., Zhang, L., Kobayashi, M., Copeland, N. G., Jenkins, N. A., and Rosenberg, R. D. (1999) Multiple isoforms of heparan sulfate D-glucosaminyl 3-O-sulfotransferase. Isolation, characterization, and expression of human cDNAs and identification of distinct genomic loci. J. Biol. Chem. 274, 5170-5184
Xia, G., Chen, J., Tiwari, V., Ju, W., Li, J. -P., Malmström, A., Shukla, D., and Liu, J. (2002) Heparan sulfate 3-O-sulfotransferase isoform 5 generates both an antithrombin-binding site and an entry receptor for herpes simplex virus, type 1. J. Biol. Chem. 277, 37912-37919
Xu, D., Tiwari, V., Xia, G., Clement, C., Shukla, D., and Liu, J. (2005) Characterization of heparan sulphate 3-O-sulphotransferase isoform 6 and its role in assisting the entry of herpes simplex virus type 1. Biochem. J. 385, 451-459
Yamada, S., Yoshida, K., Sugiura, M., Sugahara, K., Khoo, K. -H., Morris, H. R., and Dell, A. (1993) Structural studies on the bacterial lyase-resistant tetrasaccharides derived from the antithrombin III-binding site of porcine intestinal heparin. J. Biol. Chem. 268, 4780-4787
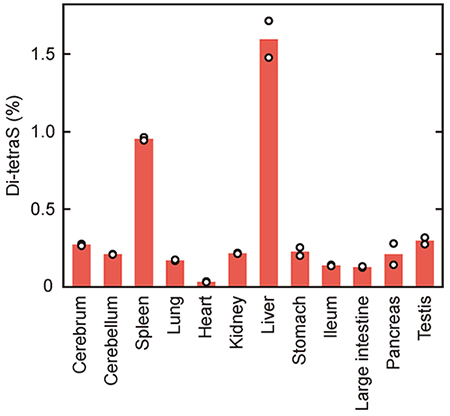
Mochizuki, H., Yoshida, K., Shibata, Y., and Kimata, K. (2008) Tetrasulfated disaccharide unit in heparan sulfate. Enzymatic formation and tissue distribution. J. Biol. Chem. 283, 31237-31245
Mochizuki, H., Yoshida, K., Gotoh, M., Sugioka, S., Kikuchi, N., Kwon, Y. -D., Tawada, A., Maeyama, K., Inaba, N., Hiruma, T., Kimata, K., and Narimatsu, H. (2003) Characterization of a heparan sulfate 3-O-sulfotransferase-5, an enzyme synthesizing a tetrasulfated disaccharide. J. Biol. Chem. 278, 26780-26787
Borjigin, J., Deng, J., Sun, X., De Jesus, M., Liu, T., and Wang, M. M. (2003) Diurnal pineal 3-O-sulphotransferase 2 expression controlled by β-adrenergic repression. J. Biol. Chem. 278, 16315-16319
Patel, V. N., Lombaert, I. M. A., Cowherd, S. N., Shworak, N. W., Xu, Y., Liu, J., and Hoffman, M. P. (2014) Hs3st3-modified heparan sulfate controls KIT+ progenitor expansion by regulating 3-O-sulfotransferases. Dev. Cell29, 662-673
Kamimura, K., Rhodes, J. M., Ueda, R., McNeely, M., Shukla, D., Kimata, K., Spear, P. G., Shworak, N. W., and Nakato, H. (2004) Regulation of Notch signaling by Drosophila heparan sulfate 3-O sulfotransferase. J. Cell Biol. 166, 1069-1079
Tecle, E., Diaz-Balzac, C. A., and Bülow, H. E. (2013) Distinct 3-O-sulfated heparan sulfate modification patterns are required for kal-1-dependent neurite branching in a context-dependent manner in Caenorhabditis elegans. G3. 3, 541-552
Denys, A., and Allain, F. (2019) The emerging roles of heparan sulfate 3-O-sulfotransferases in cancer. Front. Oncol. 9, 507
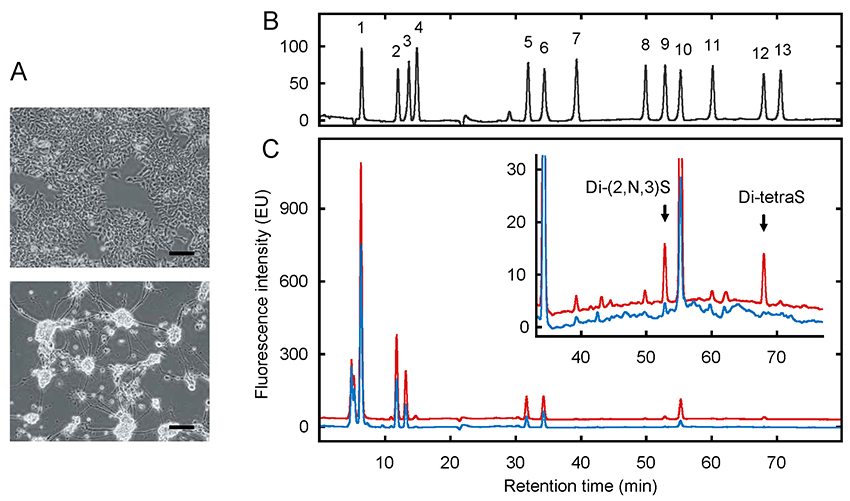
Mochizuki, H., Futatsumori, H., Suzuki, E., and Kimata, K. (2021) A quantitative method to detect non-antithrombin-binding 3-O-sulfated units in heparan sulfate. J. Biol. Chem. 296, 100115
Toyoda, H., Kinoshita-Toyoda, A., and Selleck, S. B. (2000) Structural analysis of glycosaminoglycans in Drosophila and Caenorhabditis elegans and demonstration that tout-velu, a Drosophila gene related to EXT tumor suppressors, affects heparan sulfate in vivo. J. Biol. Chem. 275, 2269-2275