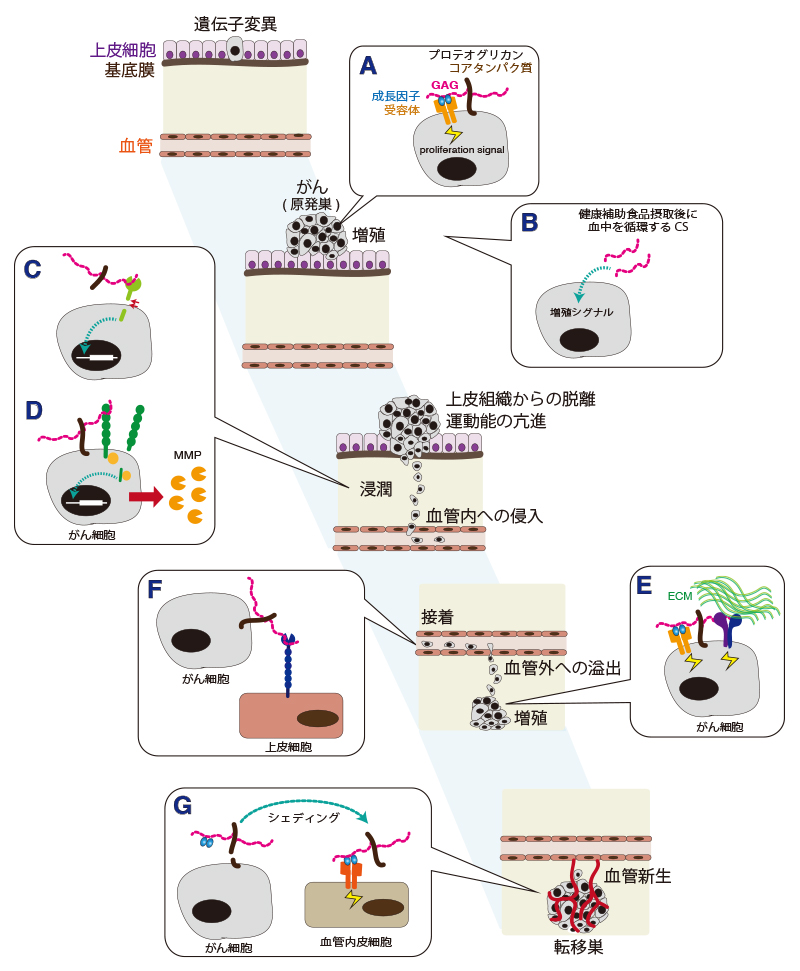
Willis, C. M., and Kluppel, M. (2014) Chondroitin sulfate-E is a negative regulator of a pro-tumorigenic Wnt/beta-catenin-Collagen 1 axis in breast cancer cells. PLoS One9, e103966
Cooney, C. A., Jousheghany, F., Yao-Borengasser, A., Phanavanh, B., Gomes, T., Kieber-Emmons, A. M., Siegel, E. R., Suva, L. J., Ferrone, S., Kieber-Emmons, T., and Monzavi-Karbassi, B. (2011) Chondroitin sulfates play a major role in breast cancer metastasis: a role for CSPG4 and CHST11 gene expression in forming surface P-selectin ligands in aggressive breast cancer cells. Breast Cancer Res13, R58
Yang, J., Price, M. A., Neudauer, C. L., Wilson, C., Ferrone, S., Xia, H., Iida, J., Simpson, M. A., and McCarthy, J. B. (2004) Melanoma chondroitin sulfate proteoglycan enhances FAK and ERK activation by distinct mechanisms. J Cell Biol165, 881-891
Yang, J., Price, M. A., Li, G. Y., Bar-Eli, M., Salgia, R., Jagedeeswaran, R., Carlson, J. H., Ferrone, S., Turley, E. A., and McCarthy, J. B. (2009) Melanoma proteoglycan modifies gene expression to stimulate tumor cell motility, growth, and epithelial-to-mesenchymal transition. Cancer Res169, 7538-7547
Lesley, J., Hyman, R., and Kincade, P. W. (1993) CD44 and its interaction with extracellular matrix. Adv Immunol54, 271-335
Kawashima, H., Atarashi, K., Hirose, M., Hirose, J., Yamada, S., Sugahara, K., and Miyasaka, M. (2002) Oversulfated chondroitin/dermatan sulfates containing GlcAbeta1/IdoAalpha1-3GalNAc(4,6-O-disulfate) interact with L- and P-selectin and chemokines. J Biol Chem277, 12921-12930
Sugahara, K. N., Hirata, T., Tanaka, T., Ogino, S., Takeda, M., Terasawa, H., Shimada, I., Tamura, J., ten Dam, G. B., van Kuppevelt, T. H., and Miyasaka, M. (2008) Chondroitin sulfate E fragments enhance CD44 cleavage and CD44-dependent motility in tumor cells. Cancer Res68, 7191-7199
Iida, J., Dorchak, J., Clancy, R., Slavik, J., Ellsworth, R., Katagiri, Y., Pugacheva, E. N., van Kuppevelt, T. H., Mural, R. J., Cutler, M. L., and Shriver, C. D. (2015) Role for chondroitin sulfate glycosaminoglycan in NEDD9-mediated breast cancer cell growth. Exp Cell Res330, 358-370
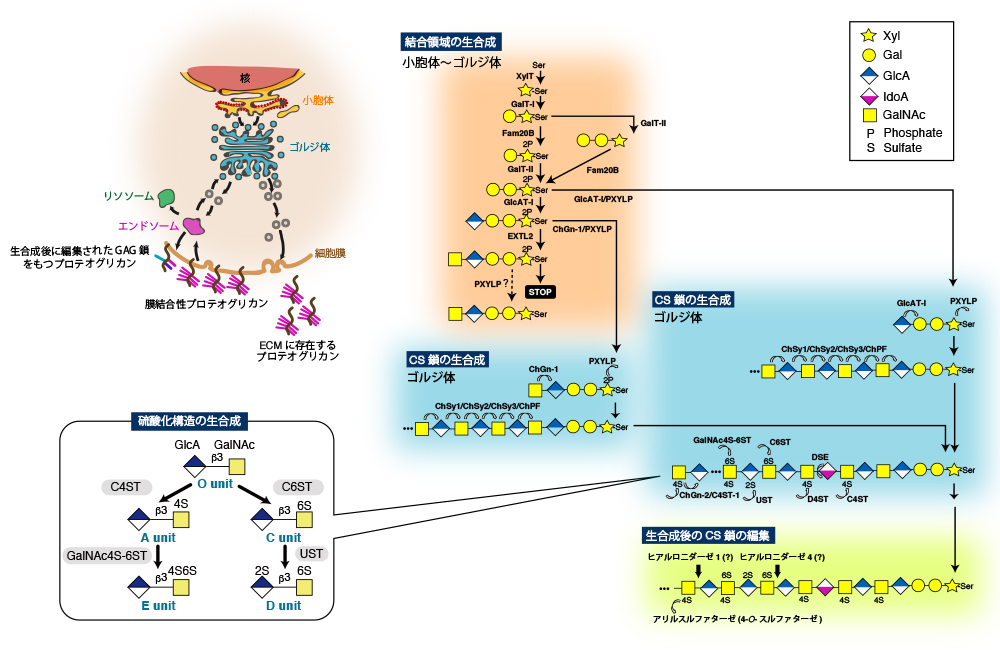
Mikami, T., and Kitagawa, H. (2013) Biosynthesis and function of chondroitin sulfate. Biochim Biophys Acta1830, 4719-4733
Nadanaka, S., and Kitagawa, H. (2008) Heparan sulphate biosynthesis and disease. J Biochem144, 7-14
Nadanaka, S., Ishida, M., Ikegami, M., and Kitagawa, H. (2008) Chondroitin 4-O-sulfotransferase-1 modulates Wnt-3a signaling through control of E disaccharide expression of chondroitin sulfate. J Biol Chem283, 27333-27343
Nadanaka, S., Kinouchi, H., Taniguchi-Morita, K., Tamura, J., and Kitagawa, H. (2011) Down-regulation of chondroitin 4-O-sulfotransferase-1 by Wnt signaling triggers diffusion of Wnt-3a. J Biol Chem286, 4199-4208
Nadanaka, S., Kinouchi, H., and Kitagawa, H. (2016) Histone deacetylase-mediated regulation of chondroitin 4-O-sulfotransferase-1 (Chst11) gene expression by Wnt/beta-catenin signaling. Biochem Biophys Res Commun480, 234-240
Hazan, R. B., Phillips, G. R., Qiao, R. F., Norton, L., and Aaronson, S. A. (2000) Exogenous expression of N-cadherin in breast cancer cells induces cell migration, invasion, and metastasis. J Cell Biol148, 779-790
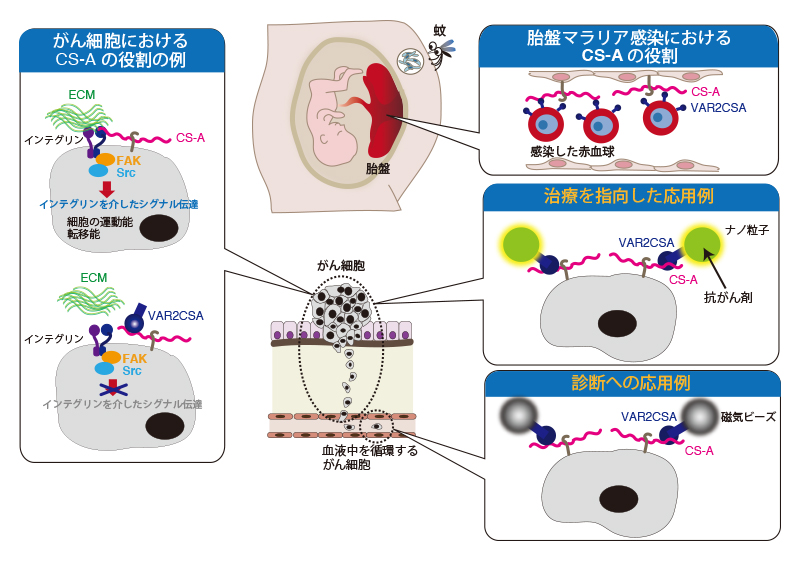
Agerbaek, M. O., Bang-Christensen, S., and Salanti, A. (2019) Fighting Cancer Using an Oncofetal Glycosaminoglycan-Binding Protein from Malaria Parasites. Trends Parasitol35, 178-181
Achur, R. N., Valiyaveettil, M., Alkhalil, A., Ockenhouse, C. F., and Gowda, D. C. (2000) Characterization of proteoglycans of human placenta and identification of unique chondroitin sulfate proteoglycans of the intervillous spaces that mediate the adherence of Plasmodium falciparum-infected erythrocytes to the placenta. J Biol Chem275, 40344-40356
Salanti, A., Clausen, T. M., Agerbaek, M. O., Al Nakouzi, N., Dahlback, M., Oo, H. Z., Lee, S., Gustavsson, T., Rich, J. R., Hedberg, B. J., Mao, Y., Barington, L., Pereira, M. A., LoBello, J., Endo, M., Fazli, L., Soden, J., Wang, C. K., Sander, A. F., Dagil, R., Thrane, S., Holst, P. J., Meng, L., Favero, F., Weiss, G. J., Nielsen, M. A., Freeth, J., Nielsen, T. O., Zaia, J., Tran, N. L., Trent, J., Babcook, J. S., Theander, T. G., Sorensen, P. H., and Daugaard, M. (2015) Targeting Human Cancer by a Glycosaminoglycan Binding Malaria Protein. Cancer Cell28, 500-514
Valente, B., Campos, P. A., do Rosario, V. E., and Silveira, H. (2010) Natural frequency of polymorphisms linked to the chondroitin 4-sulfotransferase genes and its association with placental malaria. Trans R Soc Trop Med Hyg104, 687-689
Baston-Bust, D. M., Gotte, M., Janni, W., Krussel, J. S., and Hess, A. P. (2010) Syndecan-1 knock-down in decidualized human endometrial stromal cells leads to significant changes in cytokine and angiogenic factor expression patterns. Reprod Biol Endocrinol8, 133
Clausen, T. M., Pereira, M. A., Al Nakouzi, N., Oo, H. Z., Agerbaek, M. O., Lee, S., Orum-Madsen, M. S., Kristensen, A. R., El-Naggar, A., Grandgenett, P. M., Grem, J. L., Hollingsworth, M. A., Holst, P. J., Theander, T., Sorensen, P. H., Daugaard, M., and Salanti, A. (2016) Oncofetal Chondroitin Sulfate Glycosaminoglycans Are Key Players in Integrin Signaling and Tumor Cell Motility. Mol Cancer Res14, 1288-1299
Zhang, B., Cheng, G., Zheng, M., Han, J., Wang, B., Li, M., Chen, J., Xiao, T., Zhang, J., Cai, L., Li, S., and Fan, X. (2018) Targeted delivery of doxorubicin by CSA-binding nanoparticles for choriocarcinoma treatment. Drug Deliv25, 461-471
Agerbaek, M. O., Bang-Christensen, S. R., Yang, M. H., Clausen, T. M., Pereira, M. A., Sharma, S., Ditlev, S. B., Nielsen, M. A., Choudhary, S., Gustavsson, T., Sorensen, P. H., Meyer, T., Propper, D., Shamash, J., Theander, T. G., Aicher, A., Daugaard, M., Heeschen, C., and Salanti, A. (2018) The VAR2CSA malaria protein efficiently retrieves circulating tumor cells in an EpCAM-independent manner. Nat Commun9, 3279
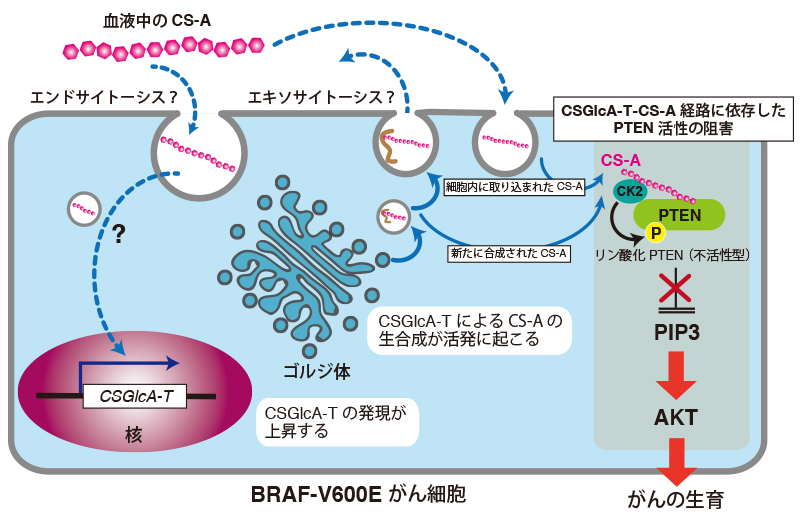
Lin, R., Xia, S., Shan, C., Chen, D., Liu, Y., Gao, X., Wang, M., Kang, H. B., Pan, Y., Liu, S., Chung, Y. R., Abdel-Wahab, O., Merghoub, T., Rossi, M., Kudchadkar, R. R., Lawson, D. H., Khuri, F. R., Lonial, S., and Chen, J. (2018) The Dietary Supplement Chondroitin-4-Sulfate Exhibits Oncogene-Specific Pro-tumor Effects on BRAF V600E Melanoma Cells. Mol Cell69, 923-937 e928
Xia, S., Lin, R., Jin, L., Zhao, L., Kang, H. B., Pan, Y., Liu, S., Qian, G., Qian, Z., Konstantakou, E., Zhang, B., Dong, J. T., Chung, Y. R., Abdel-Wahab, O., Merghoub, T., Zhou, L., Kudchadkar, R. R., Lawson, D. H., Khoury, H. J., Khuri, F. R., Boise, L. H., Lonial, S., Lee, B. H., Pollack, B. P., Arbiser, J. L., Fan, J., Lei, Q. Y., and Chen, J. (2017) Prevention of Dietary-Fat-Fueled Ketogenesis Attenuates BRAF V600E Tumor Growth. Cell Metab25, 358-373
Izumikawa, T., Koike, T., Shiozawa, S., Sugahara, K., Tamura, J., and Kitagawa, H. (2008) Identification of chondroitin sulfate glucuronyltransferase as chondroitin synthase-3 involved in chondroitin polymerization: chondroitin polymerization is achieved by multiple enzyme complexes consisting of chondroitin synthase family members. J Biol Chem283, 11396-11406
Putz, U., Howitt, J., Doan, A., Goh, C. P., Low, L. H., Silke, J., and Tan, S. S. (2012) The tumor suppressor PTEN is exported in exosomes and has phosphatase activity in recipient cells. Sci Signal5, ra70
Hopkins, B. D., Fine, B., Steinbach, N., Dendy, M., Rapp, Z., Shaw, J., Pappas, K., Yu, J. S., Hodakoski, C., Mense, S., Klein, J., Pegno, S., Sulis, M. L., Goldstein, H., Amendolara, B., Lei, L., Maurer, M., Bruce, J., Canoll, P., Hibshoosh, H., and Parsons, R. (2013) A secreted PTEN phosphatase that enters cells to alter signaling and survival. Science341, 399-402
Theocharis, A. D., Skandalis, S. S., Tzanakakis, G. N., and Karamanos, N. K. (2010) Proteoglycans in health and disease: novel roles for proteoglycans in malignancy and their pharmacological targeting. FEBS J277, 3904-3923
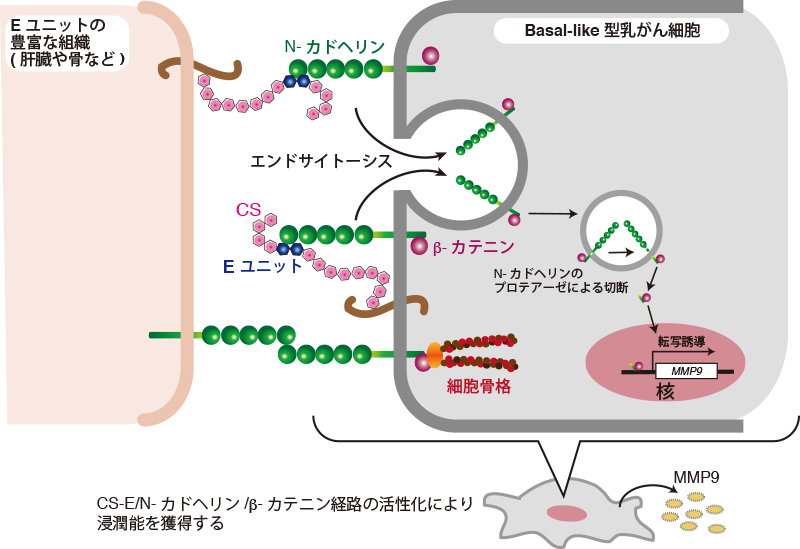
Nadanaka, S., Kinouchi, H., and Kitagawa, H. (2018) Chondroitin sulfate-mediated N-cadherin/beta-catenin signaling is associated with basal-like breast cancer cell invasion. J Biol Chem293, 444-465
Koike, T., Izumikawa, T., Tamura, J., and Kitagawa, H. (2012) Chondroitin sulfate-E fine-tunes osteoblast differentiation via ERK1/2, Smad3 and Smad1/5/8 signaling by binding to N-cadherin and cadherin-11. Biochem Biophys Res Commun420, 523-529
Nieman, M. T., Prudoff, R. S., Johnson, K. R., and Wheelock, M. J. (1999) N-cadherin promotes motility in human breast cancer cells regardless of their E-cadherin expression. J Cell Biol147, 631-644
Brown, M. S., Ye, J., Rawson, R. B., and Goldstein, J. L. (2000) Regulated intramembrane proteolysis: a control mechanism conserved from bacteria to humans. Cell100, 391-398
Reiss, K., Maretzky, T., Ludwig, A., Tousseyn, T., de Strooper, B., Hartmann, D., and Saftig, P. (2005) ADAM10 cleavage of N-cadherin and regulation of cell-cell adhesion and beta-catenin nuclear signalling. EMBO J24, 742-752
Ma, R., Feng, Y., Lin, S., Chen, J., Lin, H., Liang, X., Zheng, H., and Cai, X. (2015) Mechanisms involved in breast cancer liver metastasis. J Transl Med13, 64
Lin, R., Rosahl, T. W., Whiting, P. J., Fawcett, J. W., and Kwok, J. C. (2011) 6-Sulphated chondroitins have a positive influence on axonal regeneration. PLoS One6, e21499
Miyamoto, K., Tanaka, N., Moriguchi, K., Ueno, R., Kadomatsu, K., Kitagawa, H., and Kusunoki, S. (2014) Chondroitin 6-O-sulfate ameliorates experimental autoimmune encephalomyelitis. Glycobiology24, 469-475
Pudelko, A., Wisowski, G., Olczyk, K., and Kozma, E. M. (2019) The dual role of the glycosaminoglycan chondroitin-6-sulfate in the development, progression and metastasis of cancer. FEBS J286, 1815-1837
Herman, D., Leakey, T. I., Behrens, A., Yao-Borengasser, A., Cooney, C. A., Jousheghany, F., Phanavanh, B., Siegel, E. R., Safar, A. M., Korourian, S., Kieber-Emmons, T., and Monzavi-Karbassi, B. (2015) CHST11 gene expression and DNA methylation in breast cancer. Int J Oncol46, 1243-1251
Tan, G. K., and Tabata, Y. (2014) Chondroitin-6-sulfate attenuates inflammatory responses in murine macrophages via suppression of NF-kappaB nuclear translocation. Acta Biomater10, 2684-2692
du Souich, P., Garcia, A. G., Verges, J., and Montell, E. (2009) Immunomodulatory and anti-inflammatory effects of chondroitin sulphate. J Cell Mol Med13, 1451-1463
Engle, D. D., Tiriac, H., Rivera, K. D., Pommier, A., Whalen, S., Oni, T. E., Alagesan, B., Lee, E. J., Yao, M. A., Lucito, M. S., Spielman, B., Da Silva, B., Schoepfer, C., Wright, K., Creighton, B., Afinowicz, L., Yu, K. H., Grutzmann, R., Aust, D., Gimotty, P. A., Pollard, K. S., Hruban, R. H., Goggins, M. G., Pilarsky, C., Park, Y., Pappin, D. J., Hollingsworth, M. A., and Tuveson, D. A. (2019) The glycan CA19-9 promotes pancreatitis and pancreatic cancer in mice. Science364, 1156-1162