Higgins, G.H. and Anderson, R.M., Experimental pathology of the liver. I. Restroration of the liver of the white rat following partial surgical removal. Arch. Pathol, 1931. 12: p. 186-202.
Miyaoka, Y., et al., Hypertrophy and unconventional cell division of hepatocytes underlie liver regeneration. Curr Biol, 2012. 22(13): p. 1166-75. doi: 10.1016/j.cub.2012.05.016.
Lin, S., et al., Distributed hepatocytes expressing telomerase repopulate the liver in homeostasis and injury. Nature, 2018. 556(7700): p. 244-248. doi: 10.1038/s41586-018-0004-7.
Katsuda, T., et al., Conversion of Terminally Committed Hepatocytes to Culturable Bipotent Progenitor Cells with Regenerative Capacity. Cell Stem Cell, 2017. 20(1): p. 41-55. doi: 10.1016/j.stem.2016.10.007.
Malik, R., et al., The role of non-parenchymal cells in liver growth. Semin Cell Dev Biol, 2002. 13(6): p. 425-31.
Dieh,l, A.M. and Rai, R., Review: regulation of liver regeneration by pro-inflammatory cytokines. J Gastroenterol Hepatol, 1996. 11(5): p. 466-70.
Sekiya, S.and Suzuki, A., Glycogen synthase kinase 3 β-dependent Snail degradation directs hepatocyte proliferation in normal liver regeneration. Proc Natl Acad Sci U S A, 2011. 108(27): p. 11175-80. doi: 10.1073/pnas.1016122108.
Ding, B.S., et al., Inductive angiocrine signals from sinusoidal endothelium are required for liver regeneration. Nature, 2010. 468(7321): p. 310-5. doi: 10.1038/nature09493.
Takase HM., et al., FGF7 is a functional niche signal required for stimulation of adult liver progenitor cells that support liver regeneration. Genes Dev, 2013. 27(2): p. 169-81. doi: 10.1101/gad.204776.112.
Izumi T., et al., Vagus-macrophage-hepatocyte link promotes post-injury liver regeneration and whole-body survival through hepatic FoxM1 activation. Nat Commun, 2018. 9(1): p. 5300. doi: 10.1038/s41467-018-07747-0.
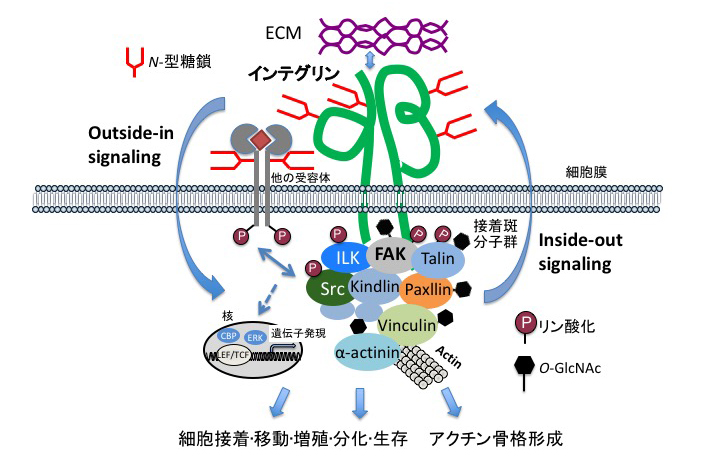
Giancotti, F.G. and Ruoslahti, E., Integrin signaling. Science, 1999. 285(5430): p. 1028-32.
Yamada, K.M. and Even-Ram, S., Integrin regulation of growth factor receptors. Nat Cell Biol, 2002. 4(4): p. E75-6.
Gu J., et al., Potential roles of N-glycosylation in cell adhesion. Glycoconj J, 2012. 29(8-9): p. 599-607. doi: 10.1007/s10719-012-9386-1.
Hang Q., et al., Integrin α5 Suppresses the Phosphorylation of Epidermal Growth Factor Receptor and Its Cellular Signaling of Cell Proliferation via N-Glycosylation. J Biol Chem, 2015. 290(49): p. 29345-60. doi: 10.1074/jbc.M115.682229.
Nagae, M., et al., Crystal structure of α5β1 integrin ectodomain: atomic details of the fibronectin receptor. J Cell Biol, 2012. 197(1): p. 131-40. doi: 10.1083/jcb.201111077.
Xu, Z., et al., O-GlcNAcylation regulates integrin-mediated cell adhesion and migration via formation of focal adhesion complexes. J Biol Chem, 2019. 294(9): p. 3117-3124. doi: 10.1074/jbc.RA118.005923.
Kikkawa, Y., et al., Transient expression of laminin alpha1 chain in regenerating murine liver: restricted localization of laminin chains and nidogen-1. Exp Cell Res, 2005. 305(1): p. 99-109.
Kikkawa, Y., et al., Laminin alpha 5 mediates ectopic adhesion of hepatocellular carcinoma through integrins and/or Lutheran/basal cell adhesion molecule. Exp Cell Res, 2008. 314(14): p. 2579-90. doi: 10.1016/j.yexcr.2008.05.021.
Tanimizu, N., et al., α1- and α5-containing laminins regulate the development of bile ducts via β1 integrin signals. J Biol Chem, 2012. 287(34): p. 28586-97. doi: 10.1074/jbc.M112.350488.
Kariya, Y., et al., N-Glycosylation of laminin-332 regulates its biological functions. A novel function of the bisecting GlcNAc. J Biol Chem, 2008. 283(48): p. 33036-45. doi: 10.1074/jbc.M804526200.
Kariya, Y., et al., Bisecting GlcNAc residues on laminin-332 down-regulate galectin-3-dependent keratinocyte motility. J Biol Chem, 2010. 285(5): p. 3330-40. doi: 10.1074/jbc.M109.038836.
Kariya, Y. and Gu, J., N-glycosylation of ß4 integrin controls the adhesion and motility of keratinocytes. PLoS One, 2011. 6(11): p. e27084. doi: 10.1371/journal.pone.0027084.
Zhao, Y., et al., Deletion of core fucosylation on alpha3beta1 integrin down-regulates its functions. J Biol Chem, 2006. 281(50): p. 38343-50.
Tamkun, J.W. and Hynes, R.O., Plasma fibronectin is synthesized and secreted by hepatocytes. J Biol Chem, 1983. 258(7): p. 4641-7.
Takamoto, M., et al., Structures of asparagine-linked oligosaccharides of human placental fibronectin. J Biochem, 1989. 105(5): p. 742-50.
Sano, K., et al., Glycosylation and ligand-binding activities of rat plasma fibronectin during liver regeneration after partial hepatectomy. Carbohydr Res, 2008. 343(13): p. 2329-35. doi: 10.1016/j.carres.2008.03.027.
Hynes, R.O., The extracellular matrix: not just pretty fibrils. Science, 2009. 326(5957): p. 1216-9. doi: 10.1126/science.1176009.
Kadota Y., et al., Mesenchymal stem cells support hepatocyte function in engineered liver grafts. Organogenesis, 2014. 10(2): p. 268-77. doi: 10.4161/org.27879.
Nadanaka, S., et al., EXTL2, a member of the EXT family of tumor suppressors, controls glycosaminoglycan biosynthesis in a xylose kinase-dependent manner. J Biol Chem, 2013. 288(13): p. 9321-33. doi: 10.1074/jbc.M112.416909.
Nadanaka, S., et al., Roles of EXTL2, a member of the EXT family of tumour suppressors, in liver injury and regeneration processes. Biochem J, 2013. 454(1): p. 133-45. doi: 10.1042/BJ20130323.
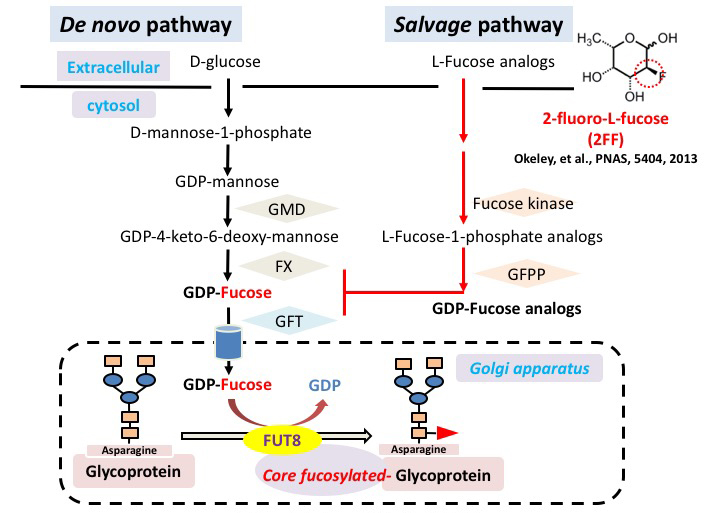
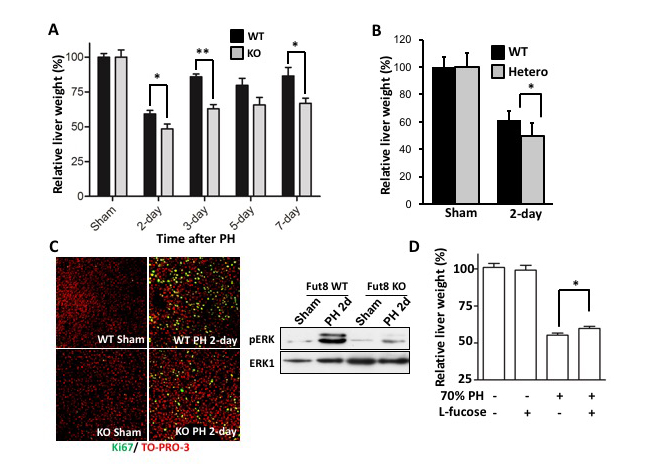
Wang, Y., et al., Loss of α1,6-fucosyltransferase suppressed liver regeneration: implication of core fucose in the regulation of growth factor receptor-mediated cellular signaling. Sci Rep, 2015. 5: p. 8264. doi: 10.1038/srep08264.
Wang, Y., et al., Loss of α1,6-fucosyltransferase inhibits chemical-induced hepatocellular carcinoma and tumorigenesis by down-regulating several cell signaling pathways. FASEB J, 2015. 29(8): p. 3217-27. doi: 10.1096/fj.15-270710.
Zhou, Y., et al., Inhibition of fucosylation by 2-fluorofucose suppresses human liver cancer HepG2 cell proliferation and migration as well as tumor formation. Sci Rep, 2017. 7(1): p. 11563. doi: 10.1038/s41598-017-11911-9.
Ng, B.G., et al., Biallelic Mutations in FUT8 Cause a Congenital Disorder of Glycosylation with Defective Fucosylation. Am J Hum Genet, 2018. 102(1): p. 188-195. doi: 10.1016/j.ajhg.2017.12.009.
Vanhooren, V., et al., Alteration in N-glycomics during mouse aging: a role for FUT8. Aging Cell, 2011. 10(6): p. 1056-66. doi: 10.1111/j.1474-9726.2011.00749.x.
Takebe, T., et al., Massive and Reproducible Production of Liver Buds Entirely from Human Pluripotent Stem Cells. Cell Rep, 2017. 21(10): p. 2661-2670. doi: 10.1016/j.celrep.2017.11.005.