次に、我々の糖鎖研究とCOPDとの関連性について経緯を述べたい。我々は以前から、N型糖鎖のコアフコース(α1-6フコースが結合した構造)を生合成する糖転移酵素であるFUT8の研究を行ってきた。まずヒト胃がん細胞株とブタ脳組織からこの酵素を均一に分離精製し、アミノ酸部分配列からcDNAクローニングを行った7,8。Fut8遺伝子のノックアウト(KO)マウスを作製したところ、7割のマウスは生後間もなく死亡したが、生存したマウスには肺気腫様の症状が見られ、マウスの呼吸機能を見たところヒトのCOPDに近い症状を呈していることがわかった9。この肺気腫症状の原因の一つは、肺胞に存在するTGF-β受容体上のコアフコースが欠損することにより、マトリックスメタロプロテアーゼの発現が高まり、肺胞を支える間質が破壊されるためであることが明らかになった9。さらに、Fut8のヘテロ欠損マウスは、COPDの危険因子であるタバコや実験的に肺気腫を惹起するエラスターゼ暴露に感受性が強く、通常のマウスでは実験的な肺気腫モデルの作製に6か月間のタバコ暴露を要するが、Fut8ヘテロ欠損マウスは3か月で同等の病態を呈する肺気腫モデルとなることがわかった10。このマウスをモデルの一つとし、ヒトのCOPDの研究を開始した。FUT8とヒトのCOPDについては、ヒトの血中FUT8の活性低下とCOPD患者の呼吸機能の低下に相関性があること11、また呼吸機能と相関するFUT8のSNPの存在が明らかになっている12。さらに最近、FUT8の発現が著しく低下する遺伝子変異によって生じるCDG (Congenital disorder of glycosylation) が見出された。このCDGは全身性に様々な症状を呈するが、共通して呼吸器症状を呈することが報告されており13、コアフコースを含む糖鎖とCOPDとの関連性がここでも示されている。
COPDは、慢性気管支炎と肺気腫と呼ばれてきた病気を包含する呼吸器疾患である。現在、日本では潜在患者をふくめると500万人以上の患者がいるとされるが、実際に治療を受けているのは30万人程度である。2012年時点の世界の死亡原因の4位であり、今後3位になると考えられている(WHO Health Statistics 2012)。COPDは生活習慣病の一つであり、危険因子は言うまでもなく喫煙であるが、PM2.5を始めとする環境因子もその原因となる。ウイルスや細菌感染により増悪することがあり、とくにインフルエンザウイルスやインフルエンザ菌、肺炎球菌による感染で増悪をもたらすことが知られている。また肺がんのリスク因子となることも論じられている。COPDの治療には、長期作用型のβ2刺激薬や長期作用型抗コリン薬、テオフィリン薬などの気管支拡張剤や吸入コルチコステロイド薬などが使われているが14、対症薬であるため、より根本的な治療薬の開発が望まれている。
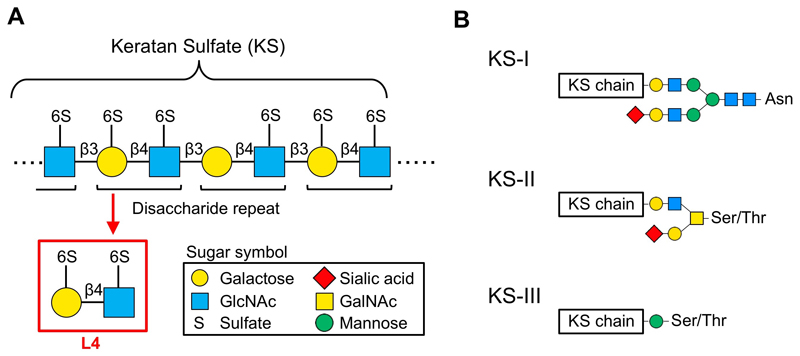
Xu, H., Kurihara, H., Ito, T., Kikuchi, H., Yoshida, K., Yamanokuchi, H., and Asari, A. (2005) The keratan sulfate disaccharide Gal(6S03) beta1,4-GlcNAc(6S03) modulates interleukin 12 production by macrophages in murine Thy-1 type autoimmune disease. J Biol Chem280, 20879-20886
Caterson, B., and Melrose, J. (2018) Keratan sulfate, a complex glycosaminoglycan with unique functional capability. Glycobiology28, 182-206
Funderburgh, J. L. (2000) Keratan sulfate: structure, biosynthesis, and function.Glycobiology10, 951-958
Akama, T. O., Nishida, K., Nakayama, J., Watanabe, H., Ozaki, K., Nakamura, T., Dota, A., Kawasaki, S., Inoue, Y., Maeda, N., Yamamoto, S., Fujiwara, T., Thonar, E. J., Shimomura, Y., Kinoshita, S., Tanigami, A., and Fukuda, M. N. (2000) Macular corneal dystrophy type I and type II are caused by distinct mutations in a new sulphotransferase gene. Nat Genet26, 237-241
Ito, Z., Sakamoto, K., Imagama, S., Matsuyama, Y., Zhang, H., Hirano, K., Ando, K., Yamashita, T., Ishiguro, N., and Kadomatsu, K. (2010) N-acetylglucosamine 6-O-sulfotransferase-1-deficient mice show better functional recovery after spinal cord injury. J Neurosci30, 5937-5947
Foyez, T., Takeda-Uchimura, Y., Ishigaki, S., Narentuya, Zhang, Z., Sobue, G., Kadomatsu, K., and Uchimura, K. (2015) Microglial keratan sulfate epitope elicits in central nervous tissues of transgenic model mice and patients with amyotrophic lateral sclerosis. Am J Pathol185, 3053-3065
Uozumi, N., Yanagidani, S., Miyoshi, E., Ihara, Y., Sakuma, T., Gao, C. X., Teshima, T., Fujii, S., Shiba, T., and Taniguchi, N. (1996) Purification and cDNA cloning of porcine brain GDP-L-Fuc:N-acetyl-beta-D-glucosaminide alpha1-->6fucosyltransferase. J Biol Chem271, 27810-27817
Yanagidani, S., Uozumi, N., Ihara, Y., Miyoshi, E., Yamaguchi, N., and Taniguchi, N. (1997) Purification and cDNA cloning of GDP-L-Fuc:N-acetyl-beta-D-glucosaminide:alpha1-6 fucosyltransferase (alpha1-6 FucT) from human gastric cancer MKN45 cells. J Biochem121, 626-632
Wang, X., Inoue, S., Gu, J., Miyoshi, E., Noda, K., Li, W., Mizuno-Horikawa, Y., Nakano, M., Asahi, M., Takahashi, M., Uozumi, N., Ihara, S., Lee, S. H., Ikeda, Y., Yamaguchi, Y., Aze, Y., Tomiyama, Y., Fujii, J., Suzuki, K., Kondo, A., Shapiro, S. D., Lopez-Otin, C., Kuwaki, T., Okabe, M., Honke, K., and Taniguchi, N. (2005) Dysregulation of TGF-beta1 receptor activation leads to abnormal lung development and emphysema-like phenotype in core fucose-deficient mice. Proc Natl Acad Sci U S A102, 15791-15796
Gao, C., Maeno, T., Ota, F., Ueno, M., Korekane, H., Takamatsu, S., Shirato, K., Matsumoto, A., Kobayashi, S., Yoshida, K., Kitazume, S., Ohtsubo, K., Betsuyaku, T., and Taniguchi, N. (2012) Sensitivity of heterozygous alpha1,6-fucosyltransferase knock-out mice to cigarette smoke-induced emphysema: implication of aberrant transforming growth factor-beta signaling and matrix metalloproteinase gene expression. J Biol Chem287, 16699-16708
Kamio, K., Yoshida, T., Gao, C., Ishii, T., Ota, F., Motegi, T., Kobayashi, S., Fujinawa, R., Ohtsubo, K., Kitazume, S., Angata, T., Azuma, A., Gemma, A., Nishimura, M., Betsuyaku, T., Kida, K., and Taniguchi, N. (2012) alpha1,6-Fucosyltransferase (Fut8) is implicated in vulnerability to elastase-induced emphysema in mice and a possible non-invasive predictive marker for disease progression and exacerbations in chronic obstructive pulmonary disease (COPD). Biochem Biophys Res Commun424, 112-117
Yamada, M., Ishii, T., Ikeda, S., Naka-Mieno, M., Tanaka, N., Arai, T., Kumasaka, T., Gemma, A., Kida, K., Muramatsu, M., and Sawabe, M. (2011) Association of fucosyltransferase 8 (FUT8) polymorphism Thr267Lys with pulmonary emphysema. J Hum Genet56, 857-860
Ng, B. G., Xu, G., Chandy, N., Steyermark, J., Shinde, D. N., Radtke, K., Raymond, K., Lebrilla, C. B., AlAsmari, A., Suchy, S. F., Powis, Z., Faqeih, E. A., Berry, S. A., Kronn, D. F., and Freeze, H. H. (2018) Biallelic Mutations in FUT8 Cause a Congenital Disorder of Glycosylation with Defective Fucosylation. Am J Hum Genet102, 188-195
Riley, C. M., and Sciurba, F. C. (2019) Diagnosis and Outpatient Management of Chronic Obstructive Pulmonary Disease: A Review. JAMA321, 786-797
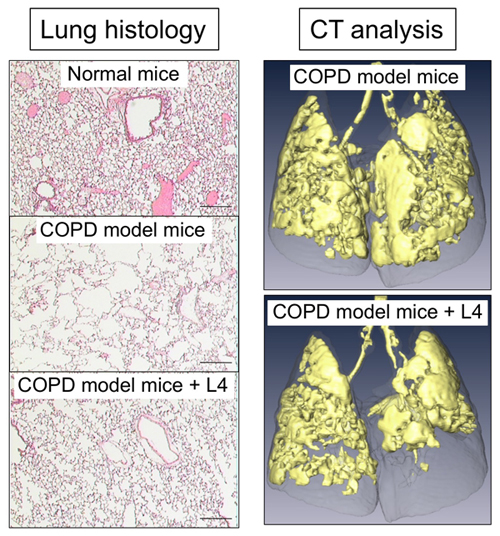
Gao, C., Fujinawa, R., Yoshida, T., Ueno, M., Ota, F., Kizuka, Y., Hirayama, T., Korekane, H., Kitazume, S., Maeno, T., Ohtsubo, K., Yoshida, K., Yamaguchi, Y., Lepenies, B., Aretz, J., Rademacher, C., Kabata, H., Hegab, A. E., Seeberger, P. H., Betsuyaku, T., Kida, K., and Taniguchi, N. (2017) A keratan sulfate disaccharide prevents inflammation and the progression of emphysema in murine models. Am J Physiol Lung Cell Mol Physiol312, L268-L276
Kobayashi, S., Fujinawa, R., Ota, F., Kobayashi, S., Angata, T., Ueno, M., Maeno, T., Kitazume, S., Yoshida, K., Ishii, T., Gao, C., Ohtsubo, K., Yamaguchi, Y., Betsuyaku, T., Kida, K., and Taniguchi, N. (2013) A single dose of lipopolysaccharide into mice with emphysema mimics human chronic obstructive pulmonary disease exacerbation as assessed by micro-computed tomography. Am J Respir Cell Mol Biol49, 971-977
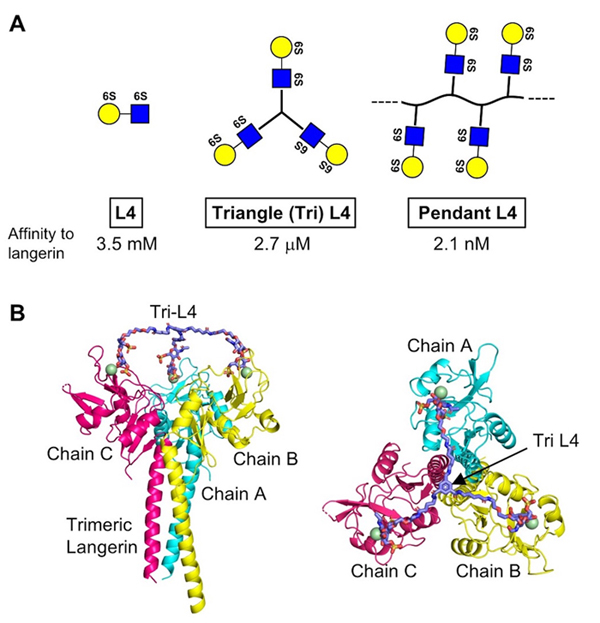
Tateno, H., Ohnishi, K., Yabe, R., Hayatsu, N., Sato, T., Takeya, M., Narimatsu, H., and Hirabayashi, J. (2010) Dual specificity of Langerin to sulfated and mannosylated glycans via a single C-type carbohydrate recognition domain. J Biol Chem 285, 6390-6400
Stambach, N. S., and Taylor, M. E. (2003) Characterization of carbohydrate recognition by langerin, a C-type lectin of Langerhans cells. Glycobiology13, 401-410
Galustian, C., Park, C. G., Chai, W., Kiso, M., Bruening, S. A., Kang, Y. S., Steinman, R. M., and Feizi, T. (2004) High and low affinity carbohydrate ligands revealed for murine SIGN-R1 by carbohydrate array and cell binding approaches, and differing specificities for SIGN-R3 and langerin. Int Immunol16, 853-866
Stoitzner, P., and Romani, N. (2011) Langerin, the "Catcher in the Rye": an important receptor for pathogens on Langerhans cells. Eur J Immunol41, 2526-2529
Bratke, K., Klug, M., Bier, A., Julius, P., Kuepper, M., Virchow, J. C., and Lommatzsch, M. (2008) Function-associated surface molecules on airway dendritic cells in cigarette smokers. Am J Respir Cell Mol Biol38, 655-660
Freeman, C. M., Martinez, F. J., Han, M. K., Ames, T. M., Chensue, S. W., Todt, J. C., Arenberg, D. A., Meldrum, C. A., Getty, C., McCloskey, L., and Curtis, J. L. (2009) Lung dendritic cell expression of maturation molecules increases with worsening chronic obstructive pulmonary disease. Am J Respir Crit Care Med180, 1179-1188
Ota, F., Hirayama, T., Kizuka, Y., Yamaguchi, Y., Fujinawa, R., Nagata, M., Ismanto, H. S., Lepenies, B., Aretz, J., Rademacher, C., Seeberger, P. H., Angata, T., Kitazume, S., Yoshida, K., Betsuyaku, T., Kida, K., Yamasaki, S., and Taniguchi, N. (2018) High affinity sugar ligands of C-type lectin receptor langerin. Biochim Biophys Acta Gen Subj1862, 1592-1601