
氏名:顧 建国
1987年に中国南通医学院を卒業。1993年に大阪大学医学部生理系博士課程修了(指導教官:生化学研究室の谷口直之教授)。その後、1993〜1997年大阪府立母子保健総合医療センター研究所の研究員(和田芳直部長); 1997〜1999年米国NIHの国立頭蓋および歯科学研究所の発生生物学部門のポスドクフェロー(Kenneth M Yamada部長);1999〜2002年大阪大学蛋白質研究所の助手(関口清俊教授)、2002年より大阪大学大学院医学系研究科生化学の助手、2004年より助教授(谷口直之教授)として、生化学・分子生物学・細胞生物学などの分野の研究に携わってきた。現在、主に細胞接着・癌転移・器官発生における糖鎖の生物学的な機能解明に関する研究を進めている。
インテグリンはすべての哺乳動物から海綿のような単純な後生動物類に至るまで、多細胞生物に広く存在する細胞外マトリックス(ECM)の受容体である 。しかし、原核生物、植物または真菌類においてはまだ同定されていない1。インテグリンは、一つのαと一つのβサブユニットからなるヘテロダイマーである。各サブユニットは大きな細胞外領域、膜貫通ドメインおよび短い細胞質部分(β4 サブユニットを除いて)を持っている。インテグリンは、細胞外ドメインでECMと結合する一方、細胞内ドメインを通じて細胞骨格制御タンパク質や非レセプター型チロシンキナーゼを含むシグナル分子と結合する (図1A)。ヒトやマウスのゲノムシーケンスに基づき、哺乳動物では、現在までに18種類のαサブユニットと8種類のβサブユニットが見つかっており、それらの組み合わせによって少なくとも24種類のインテグリンが同定されている。生体中に最も多く存在するβ1サブユニットを含むインテグリンは、図1Bに示すように12種類が同定された。各インテグリンは特異性的なリガンドを認識する。例えば、代表的な間充織型インテグリンであるα5β1は、選択的にフィブロネクチン(FN)を認識するが、上皮細胞に多く発現するインテグリンα3β1 は、基底膜の主成分であるラミニン-5(LN-5)やLN-10/11を認識する。インテグリンを介する接着は、インテグリンとそのリガンド結合に多く規定されているが、その結合の強さはインテグリンの糖鎖付加などの様々な因子によって調節されている。本稿では、インテグリンを介するシグナル伝達調節機構について、特にインテグリンα3β1とα5β1のN-結合型糖鎖の役割に焦点を絞り概説する。
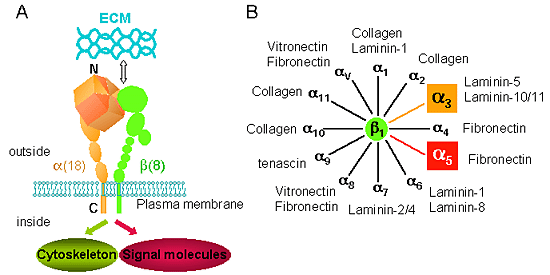
図 1 インテグリンは、一つのαと一つのβサブユニットからなるヘテロダイマーである。
各サブユニットは大きな細胞外領域、膜貫通ドメインおよび短い細胞質部分(β4サブユニットを除いて)を持っている (A)。18種類のαサブユニットが8種類のβサブユニットと組み合わせることによって現在までに24種類のインテグリンが同定されている。生体中に最も多く存在するβ1サブユニットを含むインテグリンは、これまでに12種類が同定された。各インテグリンは特異性的なリガンドを認識する (B)。
インテグリンは多細胞生物の様々な生物学のプロセスに深く関わっている。まず最も顕著な作用としては、インテグリンがECMと細胞骨格の間の物理的なリンクを提供することである。次に、インテグリンは、細胞外の情報を増殖、分化、生存などのシグナルに解読して細胞内へと(outside-in signaling)伝える役割を示している2。インテグリンを介する細胞の接着は、focal adhesion kinase (FAK)を含む様々な細胞内蛋白質のチロシンリン酸化、細胞増殖を制御するmitogen-activated protein (MAP)キナーゼの活性化、細胞生存を制御するPI 3-キナーゼ—Akt経路の活性化、さらに細胞内骨格系の再編成を制御するRhoファミリー低分子G蛋白質活性化を引き起こす (図2)。これらの経路は、細胞表面のレセプター型チロシンキナーゼを介するシグナル中にも同様に使われている。実際、インテグリンを介する細胞接着は、増殖因子受容体を介するシグナルをも制御する。その根拠としては、インテグリンを集積させた場合のみに増殖因子受容体がインテグリンの複合体に集積することである3。このように両者が物理的に結合することだけではなく、細胞内シグナルの制御にも互いに相乗効果が見られている。例として、extracellular signal-related kinase (ERK) —MAPキナーゼの活性化経路はよく研究されている4。接着系細胞が浮遊した状態では、増殖因子を添加しても瞬間的なしかも弱いERKの活性化しか起こらないが、細胞が適当なECMに接着すると、持続的かつ強いERKの活性化が起こる。一方、増殖因子はインテグリンを介するマトリックス分子のアセンブリや細胞の移動を促進する。
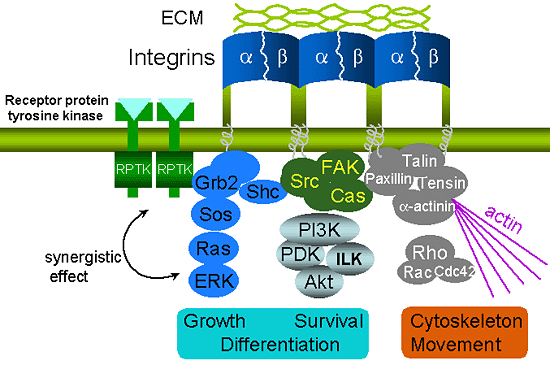
図 2 インテグリンとECMの結合は細胞内のシグナル伝達経路を活性化し、続いて細胞骨格形成・移動・増殖・分化などを調節する。
また、インテグリンは細胞表面のレセプター型チロシンキナーゼ (RPTK)と協同してこれらのシグナル活性経路を制御する。
インテグリンを介するシグナル伝達機構の研究は、これまでRGD(Arg-Gly-Asp)結合型インテグリンの一つであるα5β1とその特異的なリガンドであるFNとの相互作用の解析を中心に進められてきた。その結果、インテグリンα5β1は細胞接着・移動だけではなく、細胞の分化・生死をも制御することが明らかになった5。しかし、リガンドの特異性によって、インテグリンを介するシグナルが異なる。FNに接着した内皮細胞は、増殖因子の刺激によって増殖活性を示す。ところが、同様な細胞がLN-1に接着すると、増殖活性がなくなり、逆に増殖抑制を示す。また、LN-10/11から基底膜型インテグリンα3β1を介するシグナルは、FNからα5β1を介するシグナルと異なって、癌細胞の運動を強く促進することや6、血清飢餓によるアポトーシスを強く回避する効果を示す7。
細胞表面に発現するインテグリンは、N-グリカンの主なキャリアータンパク質である。α3β1または α5β1インテグリンのα とβサブユニットには、それぞれ14カ所と12カ所の推定されるN-結合型糖鎖付加部位を持っている。インテグリンの発現が微量でしかも精製が難しいことで、これまでにインテグリンの糖鎖構造の解析は徒労に終わる危険性がある仕事であると思われてきた。しかしながら、高橋らのグループは、異なる性質を持つHPLCカラムを利用した敏感な分析手法を開発し、胎盤からインテグリンα5β1の精製および糖鎖構造の解析に成功した8。その結果、10種類の中性糖鎖、6種類のmono-sialyl、10種類のdi-sialylおよび7種類のtri-sialyl糖鎖を合わせて35種類のオリゴ糖が同定された。また、含有の比率から見ると、中性と酸性の複合糖鎖はそれぞれ20.8%と77.7%であったのに対して、高マンノース型糖鎖は合計のたった1.5%しか含まれなかった。
一方、質量分析計を用いたヒトの尿管上皮癌細胞(HCV29)から精製したインテグリンα3β1の糖鎖構造の解析結果は、インテグリα5β1の糖鎖構造と同様に、わずかの高マンノース型糖鎖を除いて、二本鎖、三本鎖および四本鎖の複合糖鎖が大部分であることを示した9。これらの結果は、ゴルジ装置を通じてインテグリンに完全なN-結合型糖鎖が付加されることが示唆された。一般的に、適切な糖鎖の付加はタンパク質の正しいフォールディング(折りたたみ)に必要である10。実際、β1インテグリンに高マンノース型のような不完全な糖鎖が多く付加されると、これらの糖鎖が分解のシグナルとしてユビキチン分解系の複合体に認識されて、分解されてしまう11。
N-結合型糖鎖の付加は、インテグリンαとβサブユニットのアセンブリに不可欠である。実際、精製したインテグリンα5β1をN-glycosidase Fで糖鎖を切り出すと、αとβサブユニットが解離されるため、FNへの接着が完全に阻害されることが明らかとなった12。また、N-結合型糖鎖はインテグリン活性化を制御する一種のregulatorとも考えられる。ヒト線維芽細胞の培地にα-mannosidase IIのインヒビターである1-deoxymannojirimycinを添加すると、未成熟インテグリンα5β1が細胞表面に多く発現し、細胞のFNへの接着が著しく阻害された13。この結果と一致して、さらに糖鎖の非還元末端のシアル酸の有無でインテグリンα5β1の機能が大きく調節される14, 15。これらの結果は、インテグリンα5β1—FN間の結合にN-結合型糖鎖の付加だけではなく、特定の構造を持つN-結合型糖鎖の修飾も重要であることが強く示唆された。
タンパク質のN-結合型糖鎖修飾は、ゴルジ装置で様々な糖転移酵素(糖鎖遺伝子とも呼ぶ)の反応によって複雑な多様性のオリゴ糖を合成する過程である16。遺伝子導入および遺伝子ノックアウトを含む遺伝学的と生化学的アプローチを利用することが特定の糖鎖機能を知るのに有用なツールと言える17。N-アセチルグルコサミニルトランスフェラーゼIII (GnT-III)はbisecting GlcNAc糖鎖を持つユニークな分枝型糖鎖構造の合成を触媒する糖転移酵素である。bisecting GlcNAcが合成されると、ほかの分枝型糖転移酵素がこのbisected 糖鎖を基質としないため、糖鎖の伸長反応ができなくなる。したがって、GnT-IIIはN-グリカンの生合成の過程では極めて重要な糖転移酵素の一種である。このGnT-III遺伝子を導入することによって、B16マウス黒色腫細胞の肺転移能が抑制された18。その詳細なメカニズムを調べたところ、GnT-IIIの高発現細胞では、細胞表面のE-カドヘリン発現量の増加や、E-カドヘリンの下流因子であるβ-カテニンリン酸化の低下が見られた19, 20。一方、GnT-IIIを導入することで糖鎖改変されたインテグリンは、リガンドとの親和性が低下し、インテグリンを介する細胞接着・移動、また、細胞内シグナル伝達が抑制されることが明らかになった13。従って、GnT-IIIの高発現による癌転移抑制は、細胞—細胞間の接着の亢進と細胞—細胞外基質間の接着の低下という少なくとも2つのメカニズムによるものと考えられる (図3)。
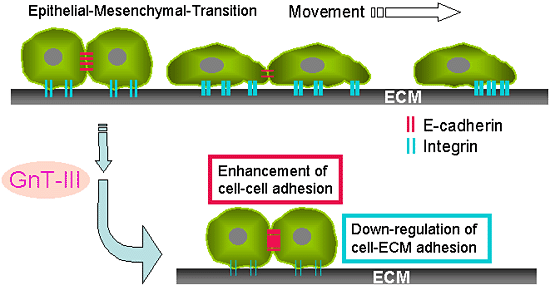
図 3 上皮から間充織の遷移(EMT)は、生体内の転移性腫瘍形成に見られる原発巣癌が浸潤性癌への進行と相関する。
GnT-IIIの高発現によるがん転移抑制には、細胞表面にE-カドヘリン発現量の上昇やβ-カテニンリン酸化の低下による細胞〜細胞間接着の亢進や、また細胞〜細胞外基質間接着の低下という少なく2つのメカニズムが関与する。
β1,6 GlcNAc分岐糖鎖構造は癌細胞侵入および癌転移の性質を持たせる可能性がある21。実際、野生型のNIH3T3細胞に比べ、発癌遺伝子Rasで形質転換したNIH3T3細胞に由来したα5β1 インテグリンには、β1,6 GlcNAcを持つ3本鎖および4本鎖が顕著に増加した22。同様に、転移性を持たない癌細胞と比較して、転移性癌細胞から精製したα3β1 インテグリンにはβ1,6 GlcNAc分岐糖鎖構造が多く見られた23。β1,6 GlcNAc分枝型糖鎖の発現と癌転移性の亢進との相関性における分子メカニズムを調べるため、Guoらは、β1,6 GlcNAc分枝型糖鎖構造をつくるGnT-VをHT1080細胞に導入した。導入された細胞では、FNによるインテグリンα5β1の過剰な集積や微小繊維へのアクチン重合が抑制され、細胞の遊走・浸潤が促進されることが明らかとなった24。このことは、インテグリンを介するECMへの細胞接着が細胞の遊走・浸潤に極めて重要であり、さらに中程度の接着は細胞の移動にとって最適な条件であるという仮説を強く裏付けることになった25。
上記のように、糖鎖リモデリングは、インテグリンの持つ細胞伸展・遊走、また細胞内シグナル伝達などの様々な生物学的な機能を制御できる。インテグリンのα とβサブユニットは、多数のN-結合型糖鎖付加サイトを持っているゆえ、インテグリン上にどのサイトが糖鎖に修飾されるか、どの糖鎖が重要であるか、または特定の糖鎖を持つインテグリンがどの分子と結合するかを同定することが必要である。したがって、このようなインテグリンのN-グリカンの機能解析は、生理的な分子間相互作用を含め、細胞内シグナリングの調節26、また、炎症、感染、癌、さらに癌転移などの病理的な過程における糖鎖の役割をより良く理解するのにとって不可欠と言える。



