|
|
Fbsファミリー:Fbsと小胞体関連分解 |
|||||||||||||||||||||||||||||
|
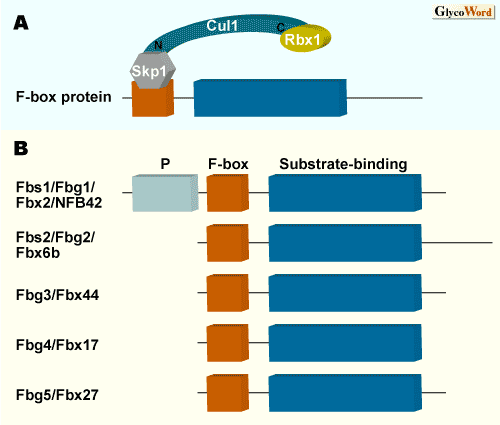
真核生物には主に2つの細胞内タンパク質分解システムがある。ひとつは非選択的分解系でバルクな分解を行なうと考えられているオートファジー系、もうひとつは個々のタンパク質寿命に関わる選択的分解を行なうユビキチン−プロテアソーム系である。後者の分解を受けるべくタンパク質の選択性の鍵を握るのがユビキチンリガーゼ(E3)と呼ばれる酵素群である。E3は哺乳類では数百種類あると考えられているが、その中で最も大きなグループがSkp1-Cul1-Rbx1/Roc1-F-boxタンパク質からなるSCF複合体と呼ばれるものである。SCF複合体を形成する4種類のタンパク質の中で、F-boxタンパク質だけが可変因子であり、これがおよそ100種類あることで大きな多様性が生まれている。F-boxタンパク質は分解を受けるタンパク質の何らかの修飾(多くはリン酸化)を感知し、標的タンパク質に結合する基質結合ユニットである。Fbs1は糖鎖(sugar chain)を認識するF-boxタンパク質ということで命名された。Fbs1のドメイン構造は図1に示すようにN末側より、Pドメイン(機能未知)、Skp1と結合するF-boxドメイン、糖鎖を認識する基質結合ドメインからなる。 |
|||||||||||||||||||||||||||||
|
|||||||||||||||||||||||||||||
|
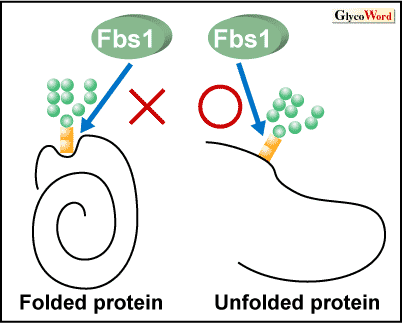
Fbs1は主に小胞体で付加されるN-結合型糖鎖の一種、高マンノース型糖鎖を認識する。Fbs1はその基質結合ドメインがX線結晶構造解析並びにNMRにより解析され、Fbs1のβサンドイッチ構造の先端にある疎水ポケットがN-結合型糖鎖の根元のキトビオース構造と結合することが明らかとなっている。糖タンパク質の根元のキトビオース構造は、通常はペプチド鎖と相互作用しているため、外側からタンパク質のような大きな分子が結合するのは難しいと考えられる。Fbs1がこのような糖鎖の根元に結合できるのは、糖タンパク質が変性することによって根元のキトビオース構造が露出する場合と考えられる。小胞体で正しい立体構造がとれなかったタンパク質は、合成系とは逆方向に小胞体から細胞質へ送り出され、ユビキチン−プロテアソーム系により分解を受けることが知られており、小胞体関連分解(ERAD)と呼ばれている。種々の研究からFbs1はこのERADのE3として働いているものと推定されている。ERAD基質は細胞質に送り出された時には変性した状態、すなわち根元のキトビオース構造が露出している状態と考えられ、Fbs1は糖鎖根元の露出を変性シグナルとして識別しているものと考えられる(図2)。 |
|||||||||||||||||||||||||||||
|
|||||||||||||||||||||||||||||
| Fbs1には基質結合ドメインの相同性の高いF-boxタンパク質が4種類存在する。これら5つのF-boxタンパク質(Fbs1/Fbg1/Fbx2/NFB42, Fbs2/Fbg2/Fbx6b, Fbg3/Fbx44, Fbg4/Fbx17, Fbg5/Fbx27)のうちFbs1以外ではFbs2の糖鎖結合能は報告されているが、他のものについてはまだ、はっきりしない。Fbs1が成体脳の神経細胞特異的に発現(Fbs1の最初に報告のあった名前はNFB42: neural F box 42 kDa であった)しているのに対し、Fbs2はその発現が多くの臓器でみられる。ERADはすべての細胞で普遍的に起こっている現象なので、Fbs2のように普遍性の高い発現様式のみられるユビキチンリガーゼが本来のERADリガーゼであるのかもしれない。Fbs1, Fbs2とも高マンノース型糖鎖を認識することは同様であるが、糖鎖に対する親和性はFbs1の方が2,3オーダー高いという特徴がある。 Fbs1,Fbs2はいずれもERAD系において構造の異常なタンパク質を小胞体から細胞質へ引きずり出す役割を担うp97/VCPと呼ばれるATPaseと相互作用することが示されている。SCFFbs1やSCFFbs2はERADにおいてp97/VCPと協調的に働いているものと考えられる。 |
|||||||||||||||||||||||||||||
| 吉田雪子(東京都臨床医学総合研究所) | |||||||||||||||||||||||||||||
|
|||||||||||||||||||||||||||||
| 2006年7月5日 | |||||||||||||||||||||||||||||
|
|
|||||||||||||||||||||||||||||
|
|||||||||||||||||||||||||||||