

|
|
摐嵔傪夘偟偨昳幙娗棟偲惗暔恑壔 |
||||||||||||||||||||||||||
 |
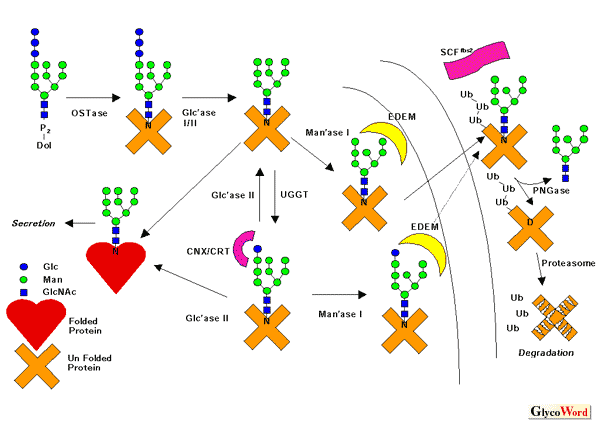
恀妀嵶朎偵偍偄偰丄彫朎懱偼暘斿宱楬傪捠傞壜梟惈偍傛傃枌僞儞僷僋幙偺崌惉偺応偱偁傞丅偙偙偱嶌傜傟傞僞儞僷僋幙偺偆偪丄婡擻揑側僞儞僷僋幙偺傒傪慖暿偟丄奺乆偺婡擻晹埵偵桝憲偡傞僔僗僥儉傪丄彫朎懱偵偍偗傞僞儞僷僋幙偺乬昳幙娗棟婡峔乭乮quality control system乯偲屇傇丅嵟嬤偺尋媶偵傛傝丄傾僗僷儔僊儞(N)宆摐嵔偺彫朎懱偵偍偗傞僾儘僙僔儞僌偼摐僞儞僷僋幙偺僼僅乕儖僨傿儞僌偺惉弉夁掱偲枾愙側娭傢傝偑偁傞偙偲偑暘偐偭偰偒偨丅懄偪丄摐嵔峔憿傪擣幆偺僞僌偲偟偰丄摐僞儞僷僋幙偺惓偟偄崅師峔憿偺妉摼傪彆挿偟偨傝丄傑偨堎忢僞儞僷僋幙偺暘夝傪懀恑偡傞丄偲偄偆栶妱偱偁傞丅偙傟傜偺婡峔偼乮1乯Glc3Man9GlcNAc2偲偄偆14摐偺僪儕僐乕儖僺儘儕儞巁偐傜僞儞僷僋幙傊偺揮堏丄乮2乯α-僌儖僐僔僟乕僛I/II偵傛傞慺憗偄扙僌儖僐僔儖壔丄乮3乯UDP-僌儖僐乕僗丗摐僞儞僷僋幙僌儖僐乕僗揮堏峺慺乮UGGT乯偵傛傞崅師峔憿晄慡僞儞僷僋幙偺慖暿偲嵞僌儖僐僔儖壔丄乮4乯僇儖僱僉僔儞乮CNX乯乛僇儖儗僥傿僉儏儕儞乮CRT乯偵傛傞儌僲僌儖僐僔儖壔摐嵔偺擣幆丄偲偄偆庤弴傪捛偭偰偄傞乮恾乯丅CNX/CRT偼偄傢備傞摐嵔埶懚揑側暘巕僔儍儁儘儞偲偟偰婡擻偟丄崅師峔憿晄慡摐僞儞僷僋幙偺嬅廤傪杊偖偲偲傕偵丄惓偟偄崅師峔憿偺妉摼傪乮捈愙揑偐丄偁傞偄偼娫愙揑偵乯懀恑偡傞丅偙偺僌儖僐乕僗偼偁傞帪娫宱偮偲α-僌儖僐僔僟乕僛II偵傛偭偰奜偝傟丄傑偨UGGT偺怰敾傪偆偗偰晄椙昳偲敾抐偝傟傟偽嵞傃僌儖僐乕僗偑晅壛偝傟偰CNX/CRT偺擣幆偺僒僀僋儖偵擖傞乮乬僇儖僱僉僔儞僒僀僋儖乭乯丅 偙傟傜偺斀墳偼偍偍傓偹恀妀嵶朎偵晛曊揑偵懚嵼偡傞偑丄婔偮偐偺椺奜偑偁傞丅乮a乯僩儕僷僲僜乕儅尨拵偼丄庬偵傛偭偰Man6GlcNAc2, Man7GlcNAc2, Man9GlcNAc2偲偄偭偨僌儖僐僔儖壔偝傟偰偄側偄僆儕僑摐傪揮堏偡傞丅偦傟屘丄UGGT偵傛傞嵞僌儖僐僔儖壔偑扙僌儖僐僔儖壔傪懸偨偢偵婲偙傞丅乮b乯弌夎峺曣乮僷儞峺曣乯偵偍偄偰偼丄CNX偺僆儖僜儘僌偵偼儌僲僌儖僐僔儖摐嵔傪擣幆偡傞擻椡偑側偔丄傑偨UGGT妶惈傕懚嵼偟側偄丅傛偭偰偄傢備傞乬僇儖僱僉僔儞僒僀僋儖乭偼懚嵼偟側偄丅扐偟慡偰偺僇價偑偦偆偱偁傞偲偄偆栿偱偼側偔丄椺偊偽幚尡儌僨儖惗暔偲偟偰抦傜傟傞暘楐峺曣偼忋婰偺僔僗僥儉傪慡偰旛偊偰偄傞傛偆偱偁傞丅 CNX/CRT偺彆偗傕嬻偟偔丄惓偟偄崅師峔憿偺妉摼偑偐側傢側偐偭偨僞儞僷僋幙偼曻抲偟偰偍偔偲撆惈偺偁傞嬅廤懱傪宍惉偡傞嫲傟傕偁傞丅偦偙偱偦傟傜偺晄梫僞儞僷僋幙偼嵶朎幙偵曻弌偝傟乮僞儞僷僋幙偺乬嵞桝憲乭乯丄僾儘僥傾僜乕儉偵傛偭偰暘夝丄彍嫀偝傟傞丅偙傟傜偺婡峔傪僞儞僷僋幙偺彫朎懱娭楢暘夝乮ER associated degradation: ERAD乯偲偄偆偑丄嵟嬤尒偄弌偝傟偨EDEM(ER degradation enhancing α-mannosidase-like protein)偲屇偽傟傞儅儞僲僔僟乕僛條僞儞僷僋幙偼丄偦偺暘巕儊僇僯僘儉偺徻嵶偼晄柧偱偁傞偑丄嫲傜偔Man8B乮恾乯偺傛偆側摐嵔峔憿傪擣幆偟偰摐僞儞僷僋幙摿堎揑偵暘夝傪懀恑偟偰偄傞傛偆偱偁傞丅偙偺EDEM暘巕傕峺曣偐傜僸僩偵帄傞傑偱丄恀妀惗暔偵峀偔曐懚偝傟偰偄傞丅
|
|||||||||||||||||||||||||
 |
||||||||||||||||||||||||||
| 丂 | ||||||||||||||||||||||||||
|
||||||||||||||||||||||||||
| 丂 |
歁擕摦暔偵偍偄偰偼丄ERAD偵傛偭偰暘夝偝傟傞僞儞僷僋幙偑嵶朎幙偵曻傝弌偝傟偨偁偲丄Fbs1/2乮Fbx2偲偦偺僷儔儘僌乯偲屇偽傟傞儐價僉僠儞儕僈乕僛偵傛偭偰摐嵔晹暘偑擣幆偝傟丄僾儘僥傾僜乕儉偵傛傞暘夝偺擣幆僞僌偱偁傞儐價僉僠儞壔偑婲偙傞丅偙偺宱楬偵娭偟偰偼柧妋側僆儖僜儘僌偑壓摍恀妀嵶朎偵偼尒摉偨傜偢丄摨條側婡峔偑暿偺暘巕偵傛偭偰扴傢傟傞偺偐丄偦傟偲傕崅摍摦暔摿堎揑側婡峔側偺偐偼崱屻偺夝愅偑懸偨傟傞偲偙傠偱偁傞丅N宆摐嵔偼僾儘僥傾僜乕儉偵傛傞暘夝傪偆偗傞慜偵PNGase偵傛偭偰摐嵔偺扙棧傪庴偗傞丅偙偺峺慺偼恀妀嵶朎偵峀偔曐懚偝傟偰偄傞丅偨偩偟摐嵔偺扙棧斀墳偺屻丄偦偺戙幱偺宱楬偼歁擕摦暔偲弌夎峺曣偱偼戝偒偔堎側傞丅戙幱偺暘巕儊僇僯僘儉偵偮偄偰偼偳偺惗暔宯偵偮偄偰傕杦偳晄柧側偺偑尰忬偱偁傞丅 |
|||||||||||||||||||||||||
| 楅栘丂嫥乮戝嶃戝妛戝妛堾堛妛宯尋媶壢惗壔妛嫵幒乯 | ||||||||||||||||||||||||||
|
||||||||||||||||||||||||||
| 丂 |
丂 |
|||||||||||||||||||||||||
| 丂 | ||||||||||||||||||||||||||
| 2004擭12寧28擔 | ||||||||||||||||||||||||||
|
|
||||||||||||||||||||||||||
|
||||||||||||||||||||||||||

