

|
|
ガレクチンによる免疫制御 |
|||||||||||||||||||||||||||||
 |
免疫機構は先天性免疫と獲得免疫からなり、協調的にかつ有効に病原体の侵入等から我々の体を防御している。様々なレクチンファミリー、例えば、セレクチンやコレクチンファミリーがこの免疫機構に関与していることが確立されている。免疫に関与しているほとんどのレクチンが膜タンパク質である。興味深い例外としてあげられるのが、この項で説明するガレクチン、そして、宿主細胞と病原菌を区別認識できるコレクチンである。 |
||||||||||||||||||||||||||||
 |
|||||||||||||||||||||||||||||
|
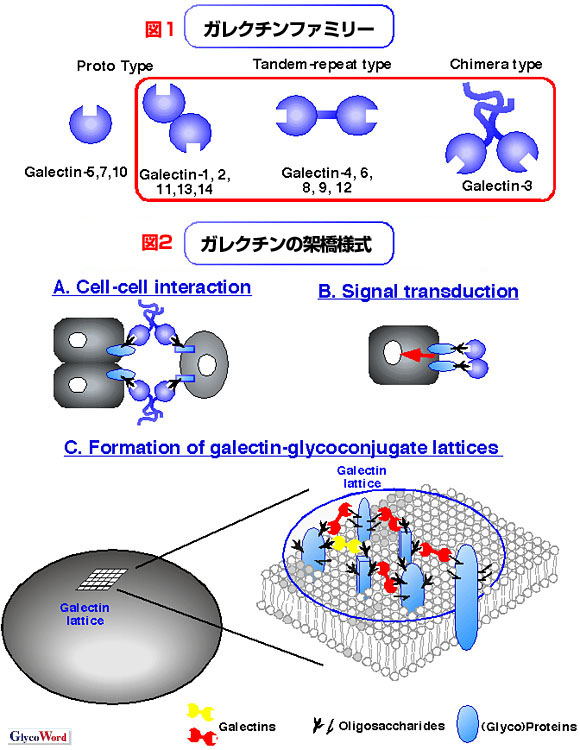
免疫反応や炎症反応の過程では、まず、白血球が炎症局所に引き寄せられる。その局所で、白血球は侵入した外来細胞(または病原体)や死細胞を貪食する。さらに、様々な因子、例えば、殺菌性の因子や免疫カスケードを始動することのできるサイトカインを分泌する。また、これらの過程の中で、個体保全のため、損傷を受けた白血球や活性化T細胞のサブセットの一部にアポトーシスが誘導される。また、炎症反応後期には、局所の修復および炎症の消散を経た炎症局所の機能的な回復が必要である。したがって、血管形成や創傷治癒が局所で誘導される。近年の報告から、ガレクチンがリガンド架橋を通して、これらの多くの過程に影響を及ぼすことが明らかにされてきている。
炎症局所や感染巣には、白血球、例えば、好中球、マクロファージやリンパ球が血流から患部に浸潤する。異なる種類の走化性因子(Chemoattractants)が白血球の局所への効率的なベクトル的移動を促進する。ガレクチン−3は単球、マクロファージに対する走化性因子であり(但し、好中球に対しての活性はない)、また、ガレクチン−9は好酸球特異的な走化性因子である。白血球の効率的な遊走には、さらに多様な細胞接着分子の調和のとれた制御が必須である。いくつかのガレクチン(例えば、ガレクチン−1,3,8,9)は細胞移動過程に関係している。興味深いことに、ガレクチンは条件により二相性の効果(接着促進効果と抗接着効果)を誘導する。ガレクチンによる接着促進は、図2Aに示したような異なる細胞上のリガンドを直接架橋することにより、または、図2Bに示したようなガレクチンによるシグナル伝達により、インテグリンのような接着分子が活性化されることにより起こる可能性が指摘されている。これに対して、いくつかの抗接着活性は白血球上のガレクチン格子が原因で誘導されるのではないかと思われる。 ガレクチン−1および−3は好中球において、侵入する病原体を殺傷できる反応性酸素中間体(reactive oxygen intermediates)を遊離する一過的な活性酸素発生(oxidative burst)を起こす。さらに、ガレクチン−3はマスト細胞の脱顆粒を誘導する。サイトカイン生成もガレクチンにより調節されたり引き起こされたりする。例えば、ガレクチン−1はTNF-α、IL-2、 IL-12、Interferon-γなどのTh1性のサイトカインの生成を阻害することにより、どちらかというと体液性免疫に関連するTh2反応にバイアスがかかるように免疫反応を誘導する。これに対し、ガレクチン−3は多様性な免疫誘導活性をもつ。すなわち、ガレクチン−3は抗炎症性サイトカインとしてIL-1産生を増強したり、白血球の浸潤を誘導する。また、ガレクチン−3はあるアレルゲン特異的T細胞や好酸球からのTh2サイトカインの一つであるIL-5の生成を特異的に阻害するが、別のTh2サイトカインであるIL-4の産生は阻害しない。また、白血球上にガレクチン格子がある場合は、T細胞レセプター依存性の免疫反応のいくつかが抑制されるとされている。 いくつかのガレクチンは、アポトーシスによる細胞死の誘導を通して、白血球のホメオスタシスに関与している。例えば、ガレクチン−1はT細胞の集団のサブセット(未分化胸腺細胞や活性化T細胞)やB細胞およびマクロファージに結合してアポトーシスを誘導し、ガレクチン−9は好酸球やいくつかのT細胞にアポプトーシスを起こす。様々な自己免疫疾患モデル動物にガレクチン−1を投入すると(ガレクチン−1のアポトーシス誘導活性によるものと考えられている)疾患が軽減することも明らかにされている。 感染症・炎症の発症中および予後、炎症局所の機能的な回復のため血管形成や創傷治癒が促進される。最近の進歩により、ガレクチン−3や−7が創傷治癒過程における再上皮細胞化(re-epithelialization)を促進することが示唆されている。ガレクチン−3はまた、毛細血管形成を促進できる。 ガレクチンは細胞外に放出され、近傍の細胞にパラクラインに結合することにより、免疫反応を調節する役割を果たす。ガレクチンリガンドである複合糖鎖は、分泌経路や細胞膜に存在する一方、ガレクチンは細胞内に存在しているので、これらのリガンドから隔離されている。重篤な感染症・炎症が起きたとき、ガレクチンは炎症や感染で損傷を受けた細胞から受動的に放出されるか、または分化したストローマ細胞、肺胞マクロファージ、慢性炎症巣や感染巣に浸潤する炎症性マクロファージから能動的に分泌される。したがって、ガレクチンが細胞から放出されるのは、我々の生体防御システムが様々な"Danger signal"を受け免疫反応を始動しなければならない時に相当する(詳細は文献参照)。したがって、ガレクチンは"Danger signal"を免疫機能に伝達することの出来る分子であり、将来、新しいサイトカイン−“レクチノサイトカイン”の代表的な分子になるべきではないかと提唱したい。 |
|||||||||||||||||||||||||||||
| 佐藤祥子(カナダ、ラバール大学医学部付属病院研究所、感染症研究所、糖質生物学研究室) | |||||||||||||||||||||||||||||
|
|||||||||||||||||||||||||||||
| 2004年1月15日 | |||||||||||||||||||||||||||||
|
|
|||||||||||||||||||||||||||||
|
|||||||||||||||||||||||||||||

