|
娻揮堏偺儊僇僯僘儉偵偼暋嶨側懡抜奒偺僗僥僢僾偑懚嵼偡傞偑丄傆偨偮偺戝偒側梫場偵暘偗偰峫偊傞昁梫偑偁傠偆丅傂偲偮偼娻嵶朎帺懱偑嬶旛偡傞埆惈搙偱偁傝丄傕偆傂偲偮偼廻庡偺柶塽擻偱偁傞丅娻嵶朎偑寣峴惈揮堏偡傞嵺偵丄寣娗撪旂嵶朎偵敪尰偡傞E-僙儗僋僠儞偲偦偺儕僈儞僪偱偁傞娻娭楢摐嵔峈尨僔傾儕儖儖僀僗X傗僔傾儕儖儖僀僗A偲偺愙拝偼嵟弶偺廳梫側僗僥僢僾偲峫偊傜傟偰偄傞丅堦曽丄帺慠柶塽宯偱戝偒側栶妱傪壥偨偡NK(natural killer)嵶朎偼峈尨偺姶嶌側偟偱庮釃嵶朎傪彎奞偡傞儕儞僷媴偱偁傞丅NK嵶朎儗僙僾僞乕偺傂偲偮偵C宆儗僋僠儞條儗僙僾僞乕偑懚嵼偡傞偑丄昗揑偲偡傞娻嵶朎偺偳偺傛偆側峔憿傪擣幆偡傞偺偐偵娭偟偰偼晄柧側揰偑懡偄丅
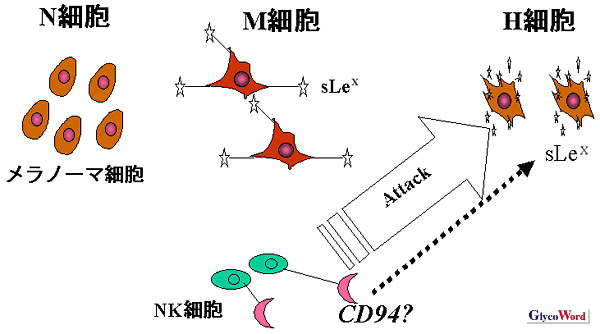
挊幰傜偑嵶朎昞柺偺僔傾儕儖儖僀僗X偺敪尰検傪曄壔偝偣丄揮堏擻偵娭偡傞専摙傪峴偭偨偲偙傠丄僔傾儕儖儖僀僗X偲NK嵶朎偲偺怴偨側娭學偑柧傜偐偵側偭偰偒偨丅傑偢丄僔傾儕儖儖僀僗X傪慡偔敪尰偟偰偄側偄B16F1儊儔僲乕儅嵶朎偵fucosyltransferase-III (FUT3) 偺cDNA傪摫擖偟偰掕忢揑敪尰僋儘乕儞姅傪嶌惢偟偨丅偙偺掕忢揑敪尰僋儘乕儞姅傪僔傾儕儖儖僀僗X偺敪尰検偵傛偭偰俁偮偺暘夋偮傑傝丄僔傾儕儖儖僀僗X傪旕忢偵嫮偔敪尰偟偰偄傞H嵶朎, 拞摍搙偵嫮偔敪尰偟偰偄傞M嵶朎, 慡偔敪尰偟偰偄側偄N嵶朎偺暘夋偵暘偗偨丅偙偺俁偮偺嵶朎姅傪C57BL/6儅僂僗旜惷柆偐傜拲幩偟偰宍惉偝傟傞攛揮堏寢愡偺悢偼M亜N=H偺弴偱懡偐偭偨丅僔傾儕儖儖僀僗X傪嵟傕懡偔敪尰偟偰偄傞H嵶朎偼攛寣娗偺拞偱傾億僩乕僔僗偵娮傝丄傎偲傫偳揮堏寢愡傪宍惉偱偒側偐偭偨丅師偵庬乆偺柶塽晄慡儅僂僗傪梡偄偰摨條偺揮堏幚尡傪偍偙側偭偨偲偙傠丄NK妶惈傪寚偔beige 儅僂僗偍傛傃峈傾僔傾儘GM1峈懱偱NK妶惈傪幐妶偝偣偨儅僂僗偱偼丄揮堏憙偺悢偼H亜M亜N偺弴偵側偭偨丅偮傑傝丄僔傾儕儖儖僀僗X偺夁忚敪尰偼寣娗撪旂嵶朎偲偺愙拝擻傪槾恑偝偣傞偑丄偦偺堦曽偱NK嵶朎偺嵶朎彎奞惈傪庝婲偝偣傞偙偲偑帵嵈偝傟偨丅
偝傜偵僸僩儊儔僲乕儅嵶朎,MeWo偵偍偄偰傕摨條偺尰徾偑婲偙傞偙偲傪妋擣偟偨丅偦偙偱丄儊儔僲乕儅嵶朎傪僞乕僎僢僩偲偟丄僸僩NK嵶朎傪僄僼僃僋僞乕偲偟偨嵶朎忈奞傾僢僙僀傪峴偭偨偲偙傠丄H>M>N偺弴偱嵶朎彎奞惈偑崅搙偱偁偭偨丅偟偐傕丄偙偺嵶朎彎奞偼CD94偵懳偡傞儌僲僋儘乕僫儖峈懱傗峈僔傾儕儖儖僀僗X儌僲僋儘乕僫儖峈懱媦傃僔傾儕儖儖僀僗X僆儕僑摐偱僽儘僢僋偝傟偨丅偝傜偵丄摐嵔峔憿夝愅偺寢壥傕峫椂偡傞偲丄僔傾儕儖儖僀僗X偺拞摍搙偺敪尰乮M嵶朎乯偼崅揮堏惈偺妉摼傪堄枴偡傞偑丄崅搙偵敪尰乮H嵶朎乯偡傞偲NK嵶朎偵傛傞峌寕傪庴偗傗偡偄偲峫偊傜傟偨丅
|
|
|
| References |
(1) |
Fukuda M: Possible roles of tumor-associated carbohydrate antigens. Cancer Res. 56, 2237-2244, 1996 |
|
(2) |
Nakamori S, Kameyama M, Imaoka S, Furukawa H, Ishikawa O, Sasaki Y, Kabuto T, Iwanaga T, Matsushita Y, Irimura T: Increased expression of sialyl Lewis X antigen correlates with poor survival in patients with colorectal carcinoma: clinicopathological and immunohistochemical study. Cancer Res. 53, 3632-3637, 1993 |
|
(3) |
Ohyama C, Tsuboi S, Fukuda M: Dual roles of sialyl Lewis X oligosaccharides in tumor metastasis and rejection by natural killer cells. EMBO J. 18, 1516-1525, 1999 |
|
|
(4) |
Ohyama C, Kanto S, Kato K, Nakano O, Arai Y, Kato T, Chen S, Fukuda MN, Fukuda M: Natural killer cells attack tumor cells expressing high levels of sialyl Lewis X oligosaccharides. Proc Natl Acad Sci U S A. 99, 13789-13794, 2002 |
|
|
|
|
Links
|
|
僔傾儕儖儖僀僗X LE-A05, GP-A03, GT-D03, GG-A00, IS-A02 |
|
|
|
|
|