
Jedrzejas博士は1988年、ポーランド、KrakowのJagellonian 大学で物理学のB.A./M.Sを、1992年Cleveland州立大学/Cleveland Clinic Foundationで化学のM.S.を、1993年構造化学のPh.D.を取得した。大学院では薬物様分子と薬理効果をもたらすタンパク質との相互作用について研究した。微生物の病原性に関するポスドク研究は、アラバマ大学・バーミンガム医科歯科学校の微生物学部で完成した。1995年、同校微生物学教室の助教授に採用された。2001年、オークランド研究所の小児病院に準教授として移った。ここでは構造生物学のグループを率いて、主としてX線結晶解析法等の構造生物学のツールを使って、高分子の構造とメカニズムの研究を続けている。グラム陽性菌の病原性、主としてStreptcoccus種に焦点を置いた微生物と宿主の相互作用の構造的視点からの研究、およびBacillusとClostridium種を重点に微生物の胞子形成、発芽、伸長を起こす必須プロセスのメカニズム研究に注力している。ヒト病原性菌の毒性因子の役割と機能の解明のための幅広い研究の一環として、最近Jedrzejas博士の研究室では、Streptcoccus種のヒアルロン酸リアーゼの結晶構造を決定し、ヒアルロン酸分解のメカニズムを解明した。
多糖分解酵素
Streptococcus種のような微生物はしばしばホストの組織への侵入、特に感染症ではさらに悪化させるのに必須な組織への侵入を容易にするため、ヒアルロン酸を含むグリカンを分解する1。グリカン構造を切断する酵素のグループは多糖分解酵素と呼ばれるが、作用機作に基づいて2つのグループ:多糖リアーゼ(脱離酵素)と多糖加水分解酵素に分けられる。一般的に多糖リアーゼはβエリミネーション(脱離反応)を利用し、例えばプロトンの受理と供与(PAD)のメカニズムあるいはその改変メカニズムに基づいて、糖鎖ポリマーを分解する(詳細は後述)。多糖加水分解酵素は分解するのに加水分解の過程を利用し、例えば直接置換あるいは二重置換のメカニズムによって行う2。加水分解メカニズムは近年三次元構造解析を用いることで、比較的新しく最近になってはじめて詳しく報告がされるようになったPADメカニズムに比べて、ずっと詳細に研究が進められている2-5。しかしながら両メカニズムとも生化学的方法では長い間、研究されてきた。幾つかの多糖分解酵素の三次元構造が解明されたことで、両者の幅広い研究は著しく進展した。これらの酵素の多くは共通したコア構造のモチーフを有しており、馬蹄形の、樽様(バレル)構造からなっている。このバレルは、一方が他方に入り込んだシリンダー状または円錐状に配列した2つのαへリックスの集合または層で表わすことができる。各層のαへリックスの数は酵素により4〜7と異なる2。このようなバレルの一端はより開放されており、通常そこに多糖分解酵素の触媒機能サイトが存在する。バレルの開放端はタンパク質全体の端から端に渡る長い溝を作る能力を有する。このような溝は多糖鎖が良く納まる立体構造を持つ。溝表面は通常陽性荷電の性質を持つアミノ酸残基が並んでいるので、表面は非常に陽性荷電となる。従って酵素は多糖鎖の陰性荷電に引きつけられて結合し易くなる。糖基質が溝に結合すると、分解触媒反応が始まる4。
ヒアルロニダーゼ
ヒアルロニダーゼは主としてヒアルロン酸を分解する多糖分解酵素の特別なグループであり、最終産物として様々な長さのオリゴ糖を産生する。ヒアルロン酸は組織の細胞外基質の一成分で、 [→4)GlcUA(β1→3)GlcNAc(β1→]nの2糖繰り返し構造から成る。GlcNAcはN-アセチル-D-グルコサミンで、GlcUAはD-グルクロン酸である (Fig. 1)。ヒアルロン酸は脊椎動物を通してほとんど全ての組織に存在し、Streptococcus zooepidemicus の様なある種の微生物にも存在する。天然物から単離されたヒアルロン酸は25,000個の2糖ユニット、あるいは107 Daにも及ぶ巨大な分子量を持つ。
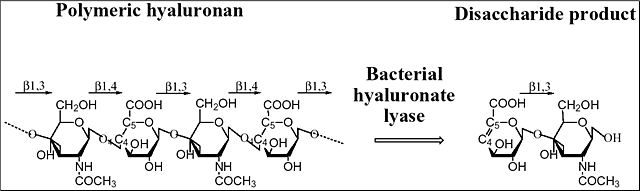
Fig. 1 ヒアルロン酸の構造と分解
ヒアルロン酸はグルクロン酸 (GluUA) とN-アセチルグルコサミン(GlcNAc)が交互に連なって構成される。ヒアルロン酸リアーゼによる分解の代表例を示す。微生物のリアーゼ作用による主たる分解生成物は、不飽和2糖である。糖鎖間のグリコシド結合も記載した。
ヒアルロニダーゼは主に3つに分類される。1)ヒアルロン酸4-グリカン加水分解酵素 (hyaluronoglucosaminidase)、例えば精巣ヒアルロニダーゼ;2) ヒアルロン酸3-グリカン加水分解酵素 (hyaluronoglucuronidase)、例えばヒルのヒアルロニダーゼ;3)ヒアルロン酸リアーゼ、 例えば微生物ヒアルロニダーゼである。最初の2つは加水分解酵素である6。ヒアルロン酸4-グリカン加水分解酵素はヒアルロン酸、コンドロイチン、ある種のコンドロイチン硫酸中のβ-N-アセチル-ヘキソサミン-(1→4)グリコシド結合を切断して、偶数単位のオリゴ糖ユニットを産生し、分解物の還元末端はN-アセチルグルコサミンとなる。ヒルのヒアルロニダーゼのようなヒアルロン酸3-グリカン加水分解酵素はヒアルロン酸のグルクロン酸結合に対して特異性があり、他の糖に対しては作用しない。最後の微生物ヒアルロニダーゼはヒアルロン酸リアーゼと呼ばれβ-エリミネーションでβ1,4結合のところでのみヒアルロン酸を分解し、しばしば2糖サイズまでの種々の長さの4,5-不飽和オリゴ糖を産生する。これは加水分解作用をしないことで、他の2グループのヒアルロニダーゼと異なっている。よく知られたヒアルロン酸リアーゼ、Streptococcus pneumoniaeとS. agalactiaeのリアーゼはヒアルロン酸を2糖に分解し、その最終産物は2-アセトアミド-2-デオキシ-3-O-(β-D-グルコ-4-エネピラノシルウロン酸)-D-グルコース(ΔDi-HA)である (Fig. 1)。同様にStreptomyces hyalurolyticusヒアルロニダーゼもエリミナーゼであるが、分解最終産物は4糖、6糖の混合物である。S. pneumoniaeやS. agalactiaeのリアーゼは、コンドロイチンや特定の位置に硫酸基が入ったコンドロイチン硫酸も分解する。これら2つのStreptpcoccusのリアーゼの三次元X線結晶構造が最近解明された2-5。酵素そのものの構造と、基質との複合体の構造が得られたことに加え、触媒メカニズムを含む生化学的特性が解析された。このような解析に立脚し、触媒メカニズムが提案された7。
決定されたS. pneumoniaeのヒアルロン酸リアーゼ (SpnHL)の三次元X線結晶構造は、約59 x 59 x 88 Åの大きさの球形タンパク質で、2つの異なった構造ドメイン、αドメインとβドメインに分かれており、両者は短いペプタイドで連結されている (Fig. 2a)5。2つのドメインは共に球状で近似サイズである。N-末端、αへリックスドメイン(αドメイン)は、活性型酵素の最初の361アミノ酸残基あるいはS. pneumoniae の全長遺伝子配列から言うとLys171からSer531を含んでいる。構造的にこのドメインは13個のαへリックスから成り、それらのグループは一回転あたりアミノ酸2〜3残基を持つコイル構造か、またはβシートや310へリックスと言った短い二次的構造から成る12個のへリックス内残基のグループで結合されている。 Asp532からSer542までの11個のリンカーアミノ酸はαドメインとβドメインを共有結合させる。C末端のβシートドメイン、βドメインはTyr543からLys889まで347個のアミノ酸から成り、5個の逆平行βシート、4つの短鎖αへリックスおよび他の二次構造要素を連結した25個のループとしてパックされた25個のβ鎖にアレンジされる。
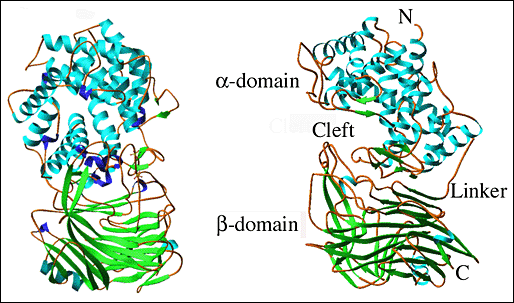
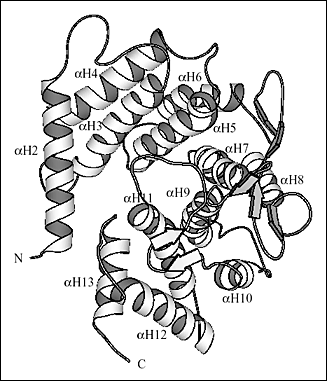
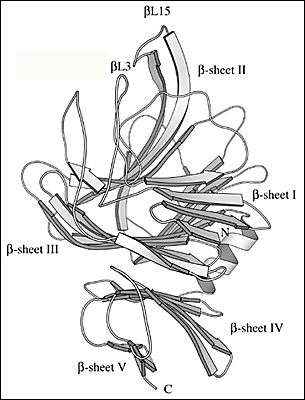
Fig. 2 S. pneumoniaeヒアルロン酸リアーゼの三次元構造
(A) 全体構造
酵素の両ドメイン、αへリックスドメイン(αドメイン、上)とβシートドメイン(βドメイン、下)を示す(左図)。酵素の90°側面図を示す(右図)。二次構造エレメント:αへリックスは青で、βシートはグリーンで、310へリックスは紫で、その他は茶で彩色。構造は1eguと命名された、www.pdb.orgのProtein Data Bankでアクセス可能な構造の座標に基づいて描かれている。
(B) αドメインの三次元構造の詳細
読者に面した端はバレルが広い側で、奧側末端は狭くなっている。へリックスはN末端から番号付けされαH1からαH13まである。バレルの狭い側の末端をブロックしているαH1は図で示されていない。
(C) βドメインの三次元構造の詳細
βドメイン全体には5つのβシートがあり、N末端から始まってβシートIからβシートVまで命名されている。αドメインに伸びている溝を作ることに係わっている2つのループはβL3とβL15(表示あり)。
αドメインの13個のαへリックス中の10個はバレル様の構造中に配列される (Fig. 2b)。バレルは一端が開放形で、他方より広がっていて不完全なものである。バレルの広がった端はβドメインを向いている。バレルの広がった側の末端はまた、隣接へリックスと連結しているループと一緒になって、SpnHL分子の中央に溝を形成している。このことが酵素の際だった特徴であり、ヒアルロン酸基質はそこに結合する。溝はαドメイン全体を横切っており、比較的まっすぐである (Fig. 2a, 2b)。ヒアルロン酸のような高分子が留まれるよう幅、深さが十分ある。βドメインは24個のβ鎖から成り5個のβシートとして配列される (Fig. 2c)。βドメイン全体は4層のサンドイッチのような形と考えられている。4つの小さなαへリックス部分はβ鎖間の幾つかの長いループに取り込まれている。つまりアミノ酸残基His578-Asn583とIle766-Ser771のβドメインから成る2つのループがあり、αドメイン中にある溝の一方の外側エッジ形成に関係している。酵素の2つのドメインはペプタイドリンカーで連結し、さらに〜3550平方Åの広さから成るドメイン間の表面インターフェイスでも相互作用する。このインターフェイスはαドメインの37残基とβドメイン34残基から形成され、中央部分はより疎水性で外側先端は親水性である。
αドメイン内にある溝はおよそ長さ30Å、深さ10Å、幅10Åである (Fig. 2a)。両端は幅広で、中央は狭い。溝幅の最短距離は約3.9Åで、αドメインのAsn290とβドメインのループの1つのAsn590の間がそれに当たる。溝は酵素がヒアルロン酸に結合し、分解するための場所として使われる5。溝に沿ったアミノ酸残基の大部分は電荷を持つか芳香族である。表面荷電の分布は溝の一方と中央部分が非常に陽性荷電になって、大きな陽性パッチを形成する (Fig. 3a)。また、3残基Glu388、Asp398とThr400は溝の片側に集まるが、そこは陽性荷電パッチ端の反対側になる。この領域には陽性に荷電した残基はなく、溝のこちら側を陰性荷電にする。このようにして3残基Glu388、Asp398とThr400は陰性パッチを形成する (Fig. 3a, 3b)。さらに3つの疎水性残基、Trp291、Trp292とPhe343が溝の一方に集まるが、それは陰性パッチのちょうど反対側になる。これが疎水性パッチを形成する (Fig. 3a, 3b)。最後に3つの触媒残基Asn349、His399とTyr408は、上記パッチの中程の、溝の最も狭いところに局在する5。
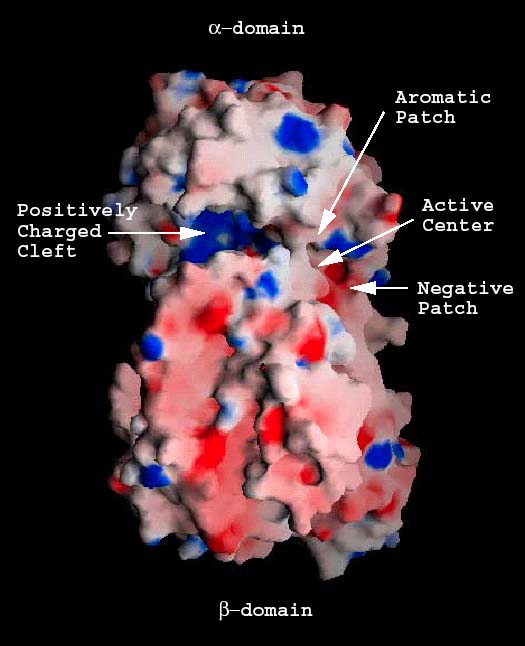
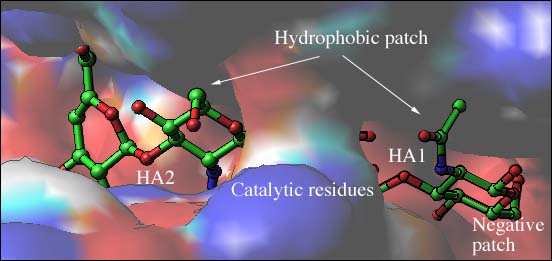
Fig. 3 S. pneumoniaeヒアルロン酸リアーゼの静電ポテンシャル
(A) 分子全体の電気ポテンシャル分布
酵素そのものの分子表面のポテンシャル分布を示す。陽性に荷電した溝(陽性パッチ、青)陰性パッチ(赤)、芳香族/疎水性パッチ(グレー)と触媒中心の残基の位置が確認できる。
(B) 結合溝の電気ポテンシャル分布
ヒアルロン酸4糖と結合した糖鎖結合溝の一部分が、触媒残基、疎水パッチ、陰性パッチに接近拡大した形で描かれている。陽性荷電領域は青で、陰性領域は赤で示した。ヒアルロン酸基質は規格に準拠して彩色した球と棒線(炭素は緑、酸素は赤、窒素は青)で表示した。HA1とHA2の2糖が描かれている。ブリッジが溝の触媒中心を超えて架かることで、分子のこの部分で溝は閉ざされている。触媒残基はブリッジ構造の下になっている。
微生物のような外来性の因子はヒアルロン酸リアーゼの酵素を使い、βエリミネーションプロセスでヒアルロン酸を分解する。逆にヒアルロン酸の動物体内での分解はホスト自身のヒアルロニダーゼによる加水分解で行われる。リアーゼ反応の正確な分子メカニズムは、比較的最近まで、すなわち微生物のヒアルロン酸リアーゼについての最初の構造に関する情報がX線結晶解析の手法によって得られるまで、ほとんど分からなかった。内在性ヒアルロニダーゼに関する構造についての情報は、今でも不明である。従ってこのプロセスの詳細はまだ不明であり、得られているものは加水分解酵素に分類される他の多糖分解酵素との比較に基づいたものである。
疎水性残基 (Trp291、Trp292とPhe343)や陰性パッチ (Glu388、Asp398とThr400)と同様、触媒残基 (Asn349、His399とTyr408)は下記1)〜3)のものを含むいくつかの研究ではっきり同定された。1) S. pneumoniaeヒアルロン酸リアーゼそのものの構造5、2) 分解産物2糖、そして4糖、6糖ヒアルロン酸ユニットとの複合体構造、3) 部位特異的突然変異体の研究である5,7。選ばれた変異体酵素がクローン化され、単離された。それらの比活性は酵素の動態パラメーターと共に決定された。変異体の活性、動態学的性質と構造の相関関係から、これらの残基が触媒反応のいろいろな部分に直接的に関与していることがはっきり確認された5,7。上述したデータの解析に基づき詳細な触媒反応メカニズムが提案された。このメカニズムは、最初に酵素によってヒアルロン酸基質からプロトンが受理され、次に酵素からβ1,4グリコシド結合の酸素に異なるプロトンが供与されることから、プロトン受理・供与 (PAD)メカニズムと命名された。分解過程は5つの別個のステップから成り立つ (Fig. 4):(1) 最初に酵素は溝の中で、主としてそこに存在する陽性荷電残基を使って陰性に荷電したヒアルロン酸基質に結合する。そして疎水性パッチを構成する残基によって、基質/酵素は触媒反応に向け正しく配置される。; (2) 最初に触媒残基 (Asp349、His399とTyr408)が関与した触媒ステップ、次いで結果的に生じるグリコシドβ1,4結合の切断と2糖分解産物の形成; (3) 酵素を元の状態に戻し次の触媒反応を進めるための微小環境中の水の分子とHis399とTyr408間の水素交換; (4) 陰性パッチ (Glu388、Asn398とThr400)の溝を利用して分解された2糖生成物が不可逆的に遊離されるステップ; (5) 残り部分の高分子量ヒアルロン酸基質を還元端の側に2糖ユニット1つ分送り出して、転位させるステップ。そのことで、β1,4グルコシド結合は次の触媒反応を進めるのに適切な位置に配置されるようになる(Fig. 4)8。これらのステップの詳細を以下に述べる。
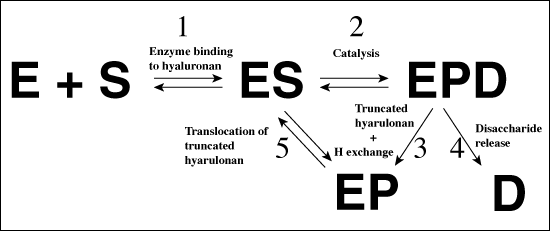
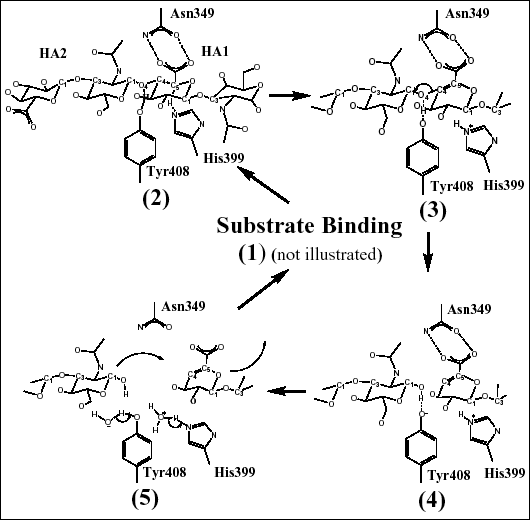
Fig. 4 ヒアルロン酸分解の概念図
(A)ヒアルロン酸リアーゼ作用の全体メカニズム
省略記号E;酵素、S;ヒアルロン酸基質、EPD;短くなったヒアルロン酸基質および2糖分解物と結合した酵素、EP;短くなったヒアルロン酸基質と結合した酵素と、酵素の活性サイトから遊離された2糖。詳細は本文参照。
(B) ヒアルロン酸のβ1,4グリコシド結合切断の触媒メカニズム
本文中で述べたヒアルロン酸分解メカニズムを説明するため、触媒残基(および基質への相対的位置)を示した。これらの残基は5段階のプロトン受理・供与メカニズムでヒアルロン酸を分解する。
ステップ1:ヒアルロン酸と酵素の結合
酵素は、αドメインを縦断する長い溝を使ってヒアルロン酸の長く伸びた鎖と結合する。溝は主として陽性荷電の性質を持ち、その伸びた形の中側に基質糖鎖が非常にうまく納まる (Fig. 5)。ヒアルロン酸は主として陰性荷電の性質を持ち、陽性荷電を有する溝と全くうまく補完し合う。酵素の溝によって作られた陽性荷電の場は、溝の中に陰性荷電の基質を導く。溝は主として酵素のαドメインの残基で作られるので、このドメインが触媒ドメインである。βシートドメインは、このドメインからの2つのループ、主としてAsn580残基が溝の形成に携わるのみで、この結合過程に際だった役割はしないようである (Fig.5)5, 8。しかしながら結晶中で不動化した酵素の構造が、この酵素での起こり得る変化や動態を完全に反映しているとは限らない。柔軟性解析が示すところによれば、酵素の2つのドメインは互いの間の位置関係について非常に大きな自由度を持つ8。それらは柔軟性のある11アミノ酸残基のリンカーでのみ結合し、またそれぞれの表面を通じて相互作用する。従って例えばαドメインとβドメインの相対的な動きは基質の触媒、溝への受け入れをこの溝の入り口を開けたり、閉めたりすることで調節させる可能性がある(詳細なディスカッションは後述)。基質への最初の結合が生じた後、ヒアルロン酸はまた、溝に一列に並んだ荷電を持つ残基と相互作用することでも溝の軸に沿って整列する5, 8。
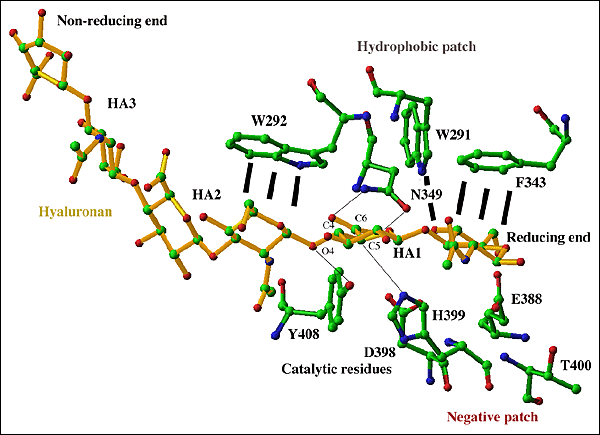
Fig. 5 S. pneumoniaeヒアルロン酸リアーゼの活性中心
6糖の糖鎖が、溝の選択された残基:触媒残基 (Asp349, His399, Phe/Tyr408)、疎水性パッチ (Trp291, Trp292, Phe343)、陰性パッチ (Glu388, Asn398, Thr400) と共に示されている。触媒グループ残基とそれらのターゲット原子の相互作用は黒線で示した。基質は黄色、酵素残基は規格に基づき彩色した(Fig. 3b参照)。還元端、非還元端の表示あり。基質の両端はもっと長く伸びていてもよい。HA1とHA2間のβ1,4グリコシド結合は酵素による触媒プロセスの結果として切断される。
基質および分解産物との酵素複合体の構造から、基質糖鎖は、糖鎖の還元末端が触媒反応が行われる溝の細い端に置かれる向きに配向される。ヒアルロン酸はこのような一方向性の結合によって、還元末端から非還元末端への分解の方向が決められる(下記参照)。触媒残基Asn349、His399とTyr408のすぐとなりに、3つの疎水性残基Trp292、Trp291とPhe343があることになる (Fig. 5)。これらの疎水性残基は基質の最初と2つ目の2糖ユニットの、糖鎖リングの疎水性の性質を使って、基質と相互作用する (Fig. 5に示したHA1とHA2の位置)。Trp292はHA2と相互作用し、一方Trp291とPhe343はHA1と相互作用する。このようにして3つの残基が基質とその化学残基を触媒反応に最も適した位置に正確に配置する (Fig. 4b)。
ステップ2:グリコシド結合の触媒的切断
3つの残基Asn349、His399とTyr408は直接的に触媒反応を起こす残基として同定された(Fig. 4b)。基質との複合体の結晶構造で可視化されたように、Asn349は直接的にHA1構成2糖のグルクロン酸部分のカルボキシル基と2箇所 (2枚歯方式) で相互作用する。この相互作用はAsn349がHA1グルクロン酸のC5の炭素からカルボキシル基の陰性荷電を引きつけることで部分的電子の貯めとして働く。このことはC5の炭素に結合している水素を酸性化させる(Fig. 4b(2))。その結果His399が塩基として働き、C5からこの水素を除去し、C5炭素の電子軌道はsp2配列に再混成される。複合体構造中で、His399はヒアルロン酸のグルクロン酸部分のC5炭素と直接的に接触する。同時にTyr408は酸として働き、グリコシド酸素O4にそのOH水素を供給し、ヒアルロン酸のHA1とHA2の2糖間のβ1,4共有グリコシド結合の切断を誘導する(Fig. 4b(3))。グリコシド酸素O4と共有結合していたC4炭素の電子軌道は同時にsp2配列に再混成され、二重結合がHA1構成2糖のグルクロン酸部分のC4とC5炭素の間に形成される(Fig. 4b(4))。結果として2糖生成物ができる (Fig. 4b)。
ステップ3:微小環境との水素交換
ステップ2で述べた触媒過程で、それぞれの酵素はHis399で水素を1つ取得し、Tyr408で1つ失う (Fig. 4b(5))。水素バランスは酵素が元の形に戻り、次の触媒反応に備えるために復元される必要がある。この構造の中で、規則的に並んだ水分子(例えば結晶構造の電子密度測定時に見られる水分子)あるいは酵素の他のアミノ酸残基を使った水素チャネリングについては、何の証拠もない。従って、水素については酵素の溝に存在する、特には構造化されていない水の微小環境を使って平衡化されていると考えられる。
ステップ4:2糖生成物の遊離
次の触媒反応を起こすため、酵素は生成した2糖を活性部位から遊離させる必要がある。2糖生成物に対応する溝の中のHA1のすぐ近くに、溝表面の底部分で集合した3つの陰性パッチ残基、Glu388、Asp398とThr400がある (Fig. 5)。これらの残基は、陰性荷電基質のHA1の2糖が結合する溝部分での陰性荷電形成に対応している (Fig. 5, 6a)。このきっちりと決まった位置にこのパッチがあることは、酵素機能上、偶然の出来事ではなく重要なことである。触媒反応後、ポリマーから2糖生成物が遊離される時、生成物がこの静電ポテンシャル中に直接的に置かれていることは明らかである。結果として反応生成物は非可逆的に溝からはじき出されていく。このような挙動から溝からの生成物の遊離が合理的に説明できる。陰性電荷のパッチがあることで生成物の遊離が容易になり、溝部分が空となり、酵素は次の触媒反応に備えることができる。

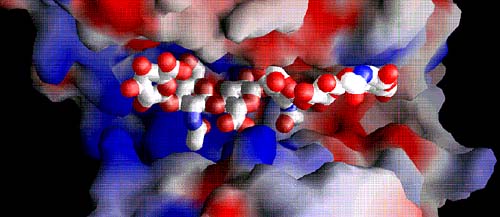
Fig. 6 6糖基質と結合した溝全体の静電ポテンシャルの分布
陽性ポテンシャルは青で、陰性は赤で示した。溝の大部分は非常に陽性荷電であり(中央と左側は陽性)、他方陰性パッチが明らかに生成物遊離端にある(右側)。S. pneumoniaeリアーゼの疎水性パッチは、基質を超えて伸びているブリッジで隠れている。S. agalactiaeリアーゼのパッチは同じ場所にある。基質は原子に対するvan der Waalsの半径を使って示されている。原子の彩色はFig. 3を参照。
(A) S. pneumoniaeリアーゼの溝に結合したヒアルロン酸のスペース充填
酵素の活性中心を覆う溝を超えて伸びるブリッジが明らかにある。溝領域の構造は酵素の閉鎖型コンフォーメーションを示している。
(B) S. agalactiaeリアーゼの溝に結合したヒアルロン酸のスペース充填
S. pneumoniaeリアーゼに比べると結合サイトの溝が広く、溝を縦断するブリッジは存在しない。溝領域の構造は酵素の開放型コンフォーメーションを示している。
ステップ5:基質の転位と酵素の連続移動的(processive)分解作用の機序
2糖分解物が遊離された後、後続分解反応の基質になり得る短くなったヒアルロン酸ポリマーが溝に残る。1つの可能性として、その基質は溝を離れ、酵素は再び触媒サイトの溝を使って新たな、あるいは同一ヒアルロン酸分子と結合する(ランダムなendo型あるいは exo型分解の結合と切断)。別の可能性として、溝に既に結合している分子がそのままそこに留まり、それから還元末端の方向に2糖ユニット分だけ、次段階の触媒反応に向けた転位が行われる(酵素により引き続き順次切り出されるexo型分解形式による結合および不飽和ヒアルロン酸2糖の切断)。S. agalactiaeヒアルロン酸リアーゼで得られた生化学的な証拠によって後者が正しいことが示された。S. pneumoniae酵素はその構造(Fig. 7)と機能的特徴、そしてアミノ酸配列がS. agalactiae酵素に似ていることから4、この酵素が同じように挙動すると考えることは妥当と思われる。この連続移動的分解過程の作用機序では、酵素は最初にヒアルロン酸にランダムに結合し、次いでポリマーを2つに切断するが(ランダムなendo型分解あるいはexo型分解)(Fig. 4a、ステップ1-5)、2糖生成物のとして上述したように還元末端側の生成物が溝を離れる。残った基質は2糖ユニット分だけ溝中で転位する。従って触媒反応は、糖鎖が十分に分解されるまで2糖を作り続ける(exo型連続移動的分解)(Fig. 4a、各ステップ2-5)。
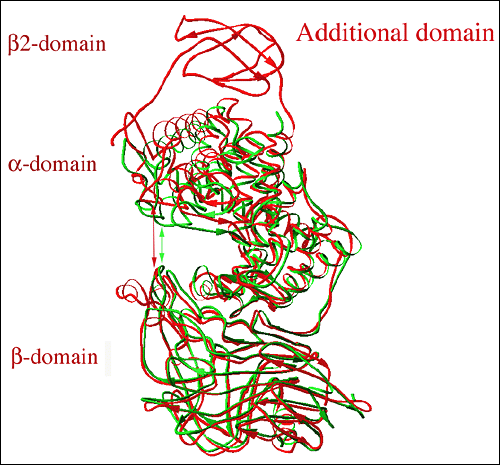
Fig. 7 Streptococcus種のヒアルロン酸リアーゼの三次元構造のアラインメント(整列)
アラインメントはS. pneumoniaeのリアーゼ(緑;www.pdb.org のpdb code: 1egu)とS. agalactiaeのリアーゼ(赤;pdb code: 1fls)の構造を基にしている。S. agalactiaeのリアーゼ構造ではもう一つβシートドメイン、すなわちβ2ドメインがN末端の部分に存在する(図の上、表示あり)。S. agalactiaeの開放型コンフォメーションの溝領域は閉鎖型コンフォメーションのS. pneumoniaeのリアーゼのものより明らかに広い。溝のおおよその幅をSagHLに対しては赤の、SpnHLに対しては緑の矢印で表示した。
高分子ヒアルロン酸が溝中で転位をなし得るために(端から順次切り出していく連続移動的機構の必須要素)、この酵素に対する幾つかの主要な要素が適所に配置されている:1) 酵素とヒアルロン酸の間の相補的境界面および基質の転位あるいは滑りが簡単にできるような直線形の結合用の溝; 2) 触媒反応後、基質が酵素と会合したままでいられるヒアルロン酸—リアーゼの正しいエネルギーのバランス、なおかつ溝に沿って動ける能力は保持している−スライドメカニズム; 3) ヒアルロン酸基質の転位/スライド過程進行中、基質の上にある酵素の疎水性パッチによって触媒サイトの場所が決められるメカニズム8,10; 4) 残った高分子ヒアルロン酸が後続の反応に向けて転位できるようスペースを作る2糖生成物を遊離するメカニズム; 5) 酵素と基質間の荷電相互作用のエネルギーから作られる転位プロセス進行用のエネルギーの存在。
ヒアルロン酸分解の方向
ヒアルロン酸分解は端から順次進む性格であることから、分解プロセスが基質の還元末端から非還元末端の方向に進むのか、逆なのかの疑問がでてきた。酵素とヒアルロン酸との複合体8および2糖生成物を含む複合体4,9の結晶構造から、分解はヒアルロン酸糖鎖の還元末端から非還元末端に向かって起こることが分かった。現在知られている構造から、4糖、6糖基質のHA1とHA2の2糖ユニット間のβ1,4グリコシド結合の切断は、糖鎖の還元末端に不飽和2糖を生じるであろうことが示されている(Fig. 5)。還元末端の基質が転位するとポリマーは新たな分解位置へ進む。そこではHA2の2糖がHA1の切断された2糖に置き換わり、HA3がHA2に置き換わっている(Fig. 4, 5)。さらにヒアルロン酸に結合した還元末端に陰性パッチがあることで、さらにこの還元末端が基質の遊離端であることを示している。
ヒアルロン酸リアーゼの柔軟性
コンピューターを使った研究で、タンパク質あるいはその一部分の運動性/動態力学と特異的機能の間の相関性が分かるようになった8。ヒアルロン酸リアーゼでは2つのタイプのタンパク質の動きがある。第一の動作はαドメイン全体と、溝から遠いβドメイン半分部分の回転あるいはひねりである。第二のものは基質—結合溝の開閉である。これはβドメインからの溝の縁を支えている2つのループ(βドメインのうち溝形成に貢献している部分)と同様、陰性パッチ残基の動きによって生じる。
第一の主たる運動によると、αドメインはヒアルロン酸結合の溝への道を開けたり閉めたりする“フタ”として働く(Fig. 2a)。この動きの中ではβドメインからはαドメインとのインターフェイスの部分も含めて残基の関与はない。また、酵素の結合溝の陽性荷電の大多数の残基は特に可動性があるが、触媒残基と陰性パッチ残基はこの酵素の柔軟性の部分にはあまり関与しない3,8。
第二の重要な動きは、基質—結合溝の開閉による触媒反応と同時に、溝に結合している基質の調整に直接的に関係している。この動きにはβドメインで溝形成に関与する2つのループだけが重要な部分をなしている。もう一つの重要な動きは触媒残基と陰性パッチの残基が関与しており(Fig. 5)、これが触媒反応と分解生成物の遊離を容易なものにしている。
S. agalactiaeヒアルロン酸リアーゼ (SagHL) の構造は最近X線結晶解析法で決定されたが4、活性サイトの幾何学的性質でも、全体構造でも肺炎球菌酵素の構造と似ていた(Fig. 7)。その構造は酵素遺伝子の完全長のAla171からIle984までを含んでいた。比較すると、SpnHLの結晶構造は肺炎球菌酵素遺伝子のLys 171からLys 984までの残基を含んでいた。配列と構造アラインメントからみると(Fig. 7)、これはSagHLのGlu249からIle984に対応している。αドメイン、ペプタイドリンカーとβドメインはSagHL中にもある。さらにS. agalactiae酵素はN-末端に主にβシート構造から成る、ここではβ2ドメインと呼ばれているAla171からVal244残基で構成される第三のドメインを持っている。付加されたβ2ドメインの存在がはっきり示されたことに加え、これら2つの肺炎球菌由来酵素の構造上の共通点からは、SpnHLの柔軟性分析で分かるようにαドメインとβドメイン間に相対的な動きがあることが示唆される(Fig. 7)。2つのドメインはお互いに離れあっているが、これはリンカーペプタイドがちょうつがいとして助けることで成り得ている。従って酵素の基質−結合溝はSpnHL構造のものよりも幅が約7Å広い(Fig. 6b, 7)。結果として触媒残基もまたお互いにかなり離れている。
SagHLアミノ酸配列では、SpnHLに比べて6個の挿入があり、削除されたものはない。全部で20残基が挿入されている。全ての挿入が分子表面で生じ、特に6個中の5個は溝の近くに固まって起きている。このような挙動はホストの免疫系を回避するために、ヒアルロン酸リアーゼ配列のバリエーションがあることを思い起させる。SagHLの溝は長さ50Å、深さ10Å、幅〜20Å(SpnHLの場合よりも〜7Å広い)である(Fig. 6b, 7)。SagHL酵素の溝構造にはSpnHL酵素の場合のような、触媒残基が局在する溝を覆うブリッジはない(Fig. 6a, 7)。SagHLでは溝の幅が広く、ブリッジがないことはヒアルロン酸リアーゼの柔軟性を反映しているらしい3,8。SagHL構造は酵素の“開放型コンフォメーション”を、SpnHL構造は酵素の“閉鎖型コンフォメーション”を示している。SpnHLに比べ、溝に沿った残基は非常に保存されており、溝表面はより荷電残基に曝され、溝形成に関与しているβドメインの残基がより多くある。SagHL における活性中心残基はSpnHL の配列と構造中に保存されている。SagHL の触媒残基はAsn429、His429とTyr488、疎水性パッチ残基はTrp371、Trp372とPhe423、陰性パッチ残基はGlu468、Asp478とThr480を含んでいる(Fig. 6b, 8)4。これらの残基は構造的にSpnHLの相同残基と非常に接近して並んでいる。さらに触媒残基は全ての既知の微生物ヒアルロン酸リアーゼの配列の中に完全に保存されている。予想されたようにSagHLの触媒残基の突然変異体は酵素活性が全くなかった。
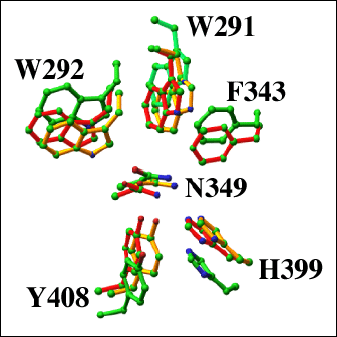
Fig. 8 各種リアーゼの触媒溝残基の構造アラインメント
S. pneumoniae(赤;pdb code: 1egu)、S. agalactiae(緑;pdb code: 1fls)とF. heparinum コンドロイチン ACリアーゼ(黄色;pdb code: 1cb8)の触媒残基Asn349、His399とTyr408を示した。さらに疎水性パッチ形成対応残基Trp291、Trp292とPhe343も示した。
ヒアルロン酸リアーゼ酵素の活性化
Streptococcus属、S. pneumonieと S. agalactiaeのヒアルロン酸リアーゼの配列解析および三次元構造情報から、酵素は活性のない前駆体フォームとしてこれら2つの病原体によって合成されることが示唆された。その後、シグナルセグメントが切られて活性化され、微生物の細胞外に活性型で分泌、放出される。共に活性型になる前に著しく分解を受ける。少なくとも、プロセスの一部は自己分解である。例えば未変化体の完全長SagHLは広範囲なタンパク質分解を受け、分子量118 kDa 、111 kDaと92 kDaの少なくとも3つの混合物となる。118 kDaのものは不活性型で、111 kDaと92 kDaのものは活性型である。SagHLの場合とは異なり、SpnHLでは全ての型について酵素の特徴が分かっているわけではないがその挙動はSagHLと似ている。
類似した三次元構造を持つ酵素
ヒアルロン酸リアーゼの触媒αドメインのバレルと類似した構造が、他の多糖分解酵素で見られる。以下に例を示す。Saccharomycopsis fibuligera グルコアミラーゼ (Protein Data Bank (PDB) code 1ayx)、Aspergillus awamori グルコアミラーゼ (PDB code 1dog)、
Clostridium thermocellum エンドグルカナーゼ CelA (PDB code 1cem)、Sphingomonas 種アルギン酸リアーゼA1-III(PDB code 1qaz)およびFlavobacterium heparinum コンドロイチンACリアーゼ (PDB code 1cb8)である (Fig. 9)。最初の3つの酵素、2種のグルコアミラーゼとエンドグルカナーゼは加水分解酵素であり、例えばS. fibuligera グルコアミラーゼはでんぷんを分解する(Fig. 9a)。Sphingomonas 種のアルギン酸リアーゼA1-IIIとF. heparinum コンドロイチンACリアーゼ (Fig. 9b) はβエリミネーションのメカニズムで分解し、アルギン酸リアーゼは海草およびある種の微生物のアルギン酸の分解を触媒する。コンドロイチンACリアーゼはヒアルロン酸とコンドロイチン硫酸を分解できるが、PADメカニズムによるらしい。多糖分解酵素の2つのグループ、加水分解酵素とリアーゼが高分子グリカンを異なるメカニズムで分解するのに、似たような三次元構造を使っていることは注目すべきである。従ってこれらの酵素は、多分共通の古い多糖に結合して分解する酵素から進化しているようだ。祖先の酵素から、分解メカニズムの細かい部分の違いが次第に進化し、後に加水分解酵素型とリアーゼ型に分かれた。基質特異性と分解プロセス、残基のコンフォメーション、位置関係、組織化にぴったり合う残基が、後に進化にあわせて選択されたようだ。
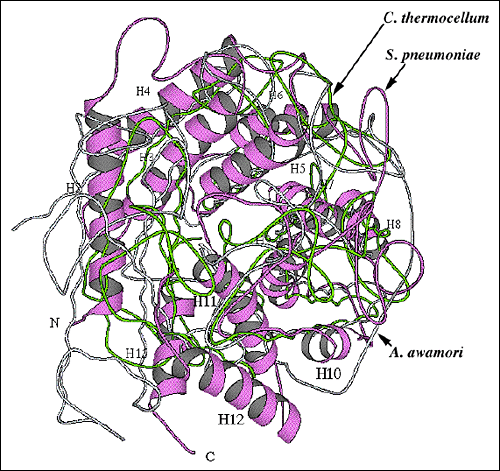
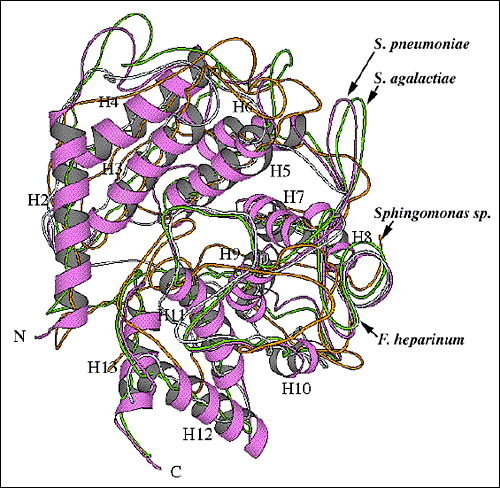
Fig. 9 各種多糖分解酵素の全体折りたたみの構造のアライメント
(A) 加水分解メカニズムを使う酵素とS. pneumoniae リアーゼとのαドメイン構造のアラインメント
S. pneumoniae リアーゼのαドメイン(紫のリボン)および2つの非常に類似した加水分解酵素C. thermocellumエンドグルカナーゼCelA(黄緑;pdb code: 1cem)とA.awamoriグルコアミラーゼ(空色;pdb code: 1dog)の構造アラインメント。加水分解酵素はαへリックスドメイン(αドメイン)のみで出来ている。構造要素が分かり易いように、S. pneumoniae ヒアルロン酸リアーゼαドメインのαへリックスに番号を付した。
(B) 他のリアーゼ酵素とS. pneumoniae リアーゼとのαドメイン構造のアラインメント
S. pneumoniae(紫のリボン)、S. agalactiae(黄緑)、F. heparinumコンドロイチン ACリアーゼ(青;pdb code:1cb8)とSphingomonas種アルギン酸リアーゼA1-III(オレンジ;pdb code: 1qaz)の構造アラインメント。S. pneumoniaeヒアルロン酸リアーゼに関してはαへリックスに番号を付した。
分解メカニズム
多糖リアーゼによる多糖の分解プロセスは、構造的および生化学的方法で検討された。最近のF. heparinum コンドロイチンACリアーゼと Sphingomonas 種アルギン酸リアーゼA1-IIIの構造的研究が、この分解プロセスに対してのさらに有益な情報を教えてくれた(Fig. 8)2,4。F. heparinumコンドロイチンACリアーゼの折りたたみ構造全体はS. pneumoniaeリアーゼと似ており、2つのドメイン、αヘリックス、βシートドメインでできている。αヘリクッスドメインは、高分子の糖鎖分子が結合して触媒反応が行われる同じ様な長い溝を支えている。コンドロイチンACリアーゼのメカニズムに対して、幾つかのシナリオが提案されたが、それらはいずれもPADメカニズムと緊密に関係している。このメカニズムには肺炎球菌のヒアルロン酸リアーゼのAsn349、His399、Tyr408に相同する残基が関係することが提案された (Fig. 8)。
ヒアルロン酸リアーゼについての現在分かっている構造情報として、S. pneumoniaeとS. agalactiae酵素本体の構造、およびヒアルロン酸分解2糖生成物と4糖、6糖基質との複合体構造がある。生化学的に得られた証拠全体と構造的データ解析をあわせて考えると、触媒のメカニズムとリアーゼの機能的性質に関しての結論がでてくる。触媒メカニズムはプロトンの受理・供与として確立されている。ヒアルロン酸分解の全過程を包含する解析から、基質の還元末端から2糖生成物が不可逆的に遊離され、ひき続いて隣接する2糖が連続移動的に分解されることが提案された。基質の結合、触媒反応のための正確な位置、触媒反応そのもの、生成物の遊離および残っている基質の転位が、ヒアルロン酸の分解全体に貢献している。さらに、触媒部分を含むヒアルロン酸結合の溝と同様に、酵素全体とそのドメインのアロステリックな挙動がヒアルロン酸リアーゼの機能を助長している。
謝辞
読者の皆さんには、このサイトの他の論文、とりわけ鈴木旺博士の“微生物のヒアルロナンリアーゼ”(ヒアルロナンリアーゼ=ヒアルロン酸リアーゼ)をお読み頂きたい。本研究はNIH (AI 44079) と国防省 (N66001-01-C-8013)の資金を得て行われたものである。



