
Stern博士は1957年ハーバード大学でB.A.を得た。彼はシアトルのワシントン大学でEdmond Fisher博士とEdwin Krebs博士の研究室で医学生として研究し、1962年M.D.を授与された。彼はイスラエルRehovothのWeizmann Institute of ScienceのUriel Littauer博士の下でポスドクとなり、米国、メリーランド州、NIH、NIDRのResearch Scientistとなった。1977年にカリフォルニア州、UCSFに移って以降、ヒアルロン酸とヒアルロニダーゼの生物学に着目している。彼はまた局公認の解剖病理学者で病理学教室で教育と診断業務に関与している。

Csóka博士は英国Newcastle大学において遺伝学でB.S.を授与され、Leicester大学において分子病理学でM.S.を授与された。彼は1998年9月ハンガリーDebrecen医科大学において細胞および分子生物学でPh.D.を授与された。彼は現在、サンフランシスコのRobert Stern博士の研究室のvisiting scientistである。ヒアルロン酸とヒアルロニダーゼに加え、加齢と幹細胞の生物学にも関心を持っている。
ヒアルロン酸のほ乳類体内での代謝は、細胞外マトリックス(ECM)構成分子としては驚くほど活発である。この過程に関与する酵素であるヒアルロニダーゼは生体内に広く分布している。これらの酵素は異なる基質特異性を有し、また、その至適pHも広い1-3。ヒアルロニダーゼ活性は、当初、ほ乳類精巣の抽出物から「拡散因子」として同定された。「ヒアルロニダーゼ」という名称は1940年にKarl Meyerによってヒアルロン酸を分解する酵素を意味する語として導入された。酵素とその反応生成物の生化学的な解析に基づき、初め彼はヒアルロニダーゼを3つの異なったクラスに分類した。遺伝子の情報の出現した現在、Meyerの分類案は驚くほど正確であったことが明らかとなっている。彼は3種の主要なヒアルロニダーゼを同定した。
1) ほ乳類タイプヒアルロニダーゼ(EC3.2.1.35)は主要な最終生成物を四糖と六糖とするエンド-β-N-アセチルヘキソサミニダーゼである。それらは加水分解活性と糖転移活性の両方を持ちa、ヒアルロン酸とコンドロイチン硫酸(CS)、特にC-4SとC-6S、そしてまた活性は低いがデルマタン硫酸(DS)も分解する。
2) 細菌性ヒアルロニダーゼ(EC4.2.99.1)はヒアルロン酸と、いろいろな程度でCSとDSを分解する。それらはエンド-β-N-アセチルヘキソサミニダーゼで主に二糖を最終生成物とするβエリミネーションによって作用するb。
3) ヒル、そのほかの寄生虫や甲殻類のヒアルロニダーゼ(EC.3.2.1.36)はエンド-β-グルクロニダーゼで最終生成物の四糖と六糖を生じる。
a ヒアルロン酸とコンドロイチン硫酸は自然界にしばしば共に存在し、お互いに結合する活性をも有する。ほ乳類ヒアルロニダーゼの糖転移活性は、試験管内でこの二種のグリコサミノグリカン鎖の間でハイブリッド分子を生み出す活性を持つ。これらが生体内で生じているのか、そして、これらのハイブリッド分子が生物学的活性を有するのかに関しては調べられていない。
b しかしながら肺炎球菌や放線菌から得たヒアルロニダーゼのように、いくつかのものは完全にヒアルロン酸のみに特異性を示す。このシリーズのS. Suzukiの総説を参照
脊椎動物組織内でのヒアルロン酸の分解は実際に次の3種の酵素の協調した反応によって起こっている。ヒアルロニダーゼと、非還元末端から糖を順次除去する2種のエキソグリコシダーゼ、すなわちβ−グルクロニダーゼとβ−N−アセチルグルコサミニダーゼである。ヒアルロニダーゼによるエンド型の開裂がエキソグリコシダーゼのために非還元末端の数を増加させる。エンド型とエキソ型の開裂反応の相対的な寄与は組織によって異なり、それらの全ヒアルロン酸分解における個々の寄与に関しては記述がない。
精巣抽出物のヒアルロニダーゼがこれまで長い間、認知され4、その性質が研究されてきた5。しかしながら、脊椎動物の臓器由来のヒアルロニダーゼは、その重要性にもかかわらず、今まで解明を拒んできた。それらは精製が難しく、非常に低濃度しか存在せず、そして、その特異的活性は非常に高いが界面活性剤や蛋白分解阻害剤非存在下では不安定である。ヒアルロニダーゼ活性の検出法の現在までの改良により6-8その精製が容易になった。
ほ乳類の生体組織から単一にまで精製された最初のほ乳類ヒアルロニダーゼはヒト血清から得られたHyal-1であった。そして遺伝子クローニング、一次構造の決定、タンパクとしての発現がなされた9。分子量57kDaで、酸性条件下で活性を持つこの酵素は49kDaの単一ポリペプチド鎖に翻訳後修飾によって8kDaの糖鎖が付加されている。この酵素は精子特異的ヒアルロニダーゼであるPH-20と40%の相同性を有する。マウスの対応する遺伝子も単離され、そのタンパクとしての発現がなされ10、ヒトの酵素と73%の相同性が観察された。
ヒト尿に有意なレベルのヒアルロニダーゼ活性が含まれることは長い間証明されてきた。ヒト尿中にはHyal-1と同一の酵素が、血清中に見つかる比活性の100倍を有して存在する。他に活性を持つ第二のバンドとしてプロセッシングを受けた45kDaのタンパクが尿中にみられる。それは、あたかも大きい方のタンパクが2カ所のエンド型蛋白分解反応によってカルボキシル末端から100アミノ酸を欠失し(Fig. 1)、二つのポリペプチド鎖がジスルフィド結合によって互いに結合しているかのようである11。この第二のバンドの活性の重要性については不明であり、この蛋白分解酵素によるプロセッシングがおこる場所が腎臓かどうかもはっきりしていない。われわれはこの酵素の第二のタイプはリソソームに存在するという予備的な証拠を持っている。
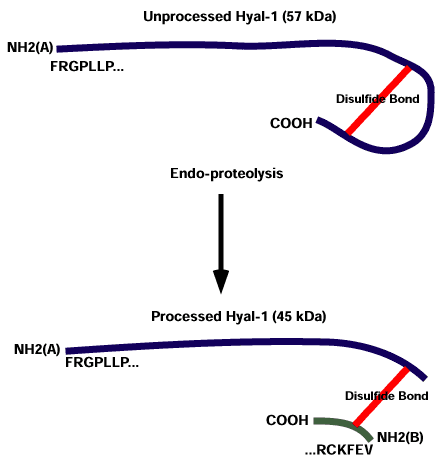
Fig. 1 Hyal-1の推定上のエンド型蛋白分解酵素によるプロセッシングの図。
プロセッシングを受けないHyal-1を上の図に示す。エンド型蛋白分解酵素によるプロセッシングの後、二つのフラグメントが生成され、二つの分離したN末端を作り出す。22アミノ酸からなるフラグメントはおそらくジスルフィド結合によりもう一方のタンパクに結合している。
ヒトHyal-1遺伝子、慣習上HYAL1と記述する、の配列がわかり、ESTデータベースの検索が可能となった。この解析に加えて、熱帯性マラリア原虫微生物の予備的なゲノムデータベース中にヒトの配列の混入が偶然に見つかったことにより、ヒト遺伝子はそれぞれが約40%の相同性を有する6個のパラロガスヒアルロニダーゼを持つことが明らかとなった。それらはヒト染色体上の3p21.3と7q31.3の2カ所に近接して連結した3連の遺伝子にグループ化されている(Fig. 2)。このような配置は太古に2回の重複からできたcluster blockが近年さらにen masse cluster-block重複が起こったためかもしれない12。
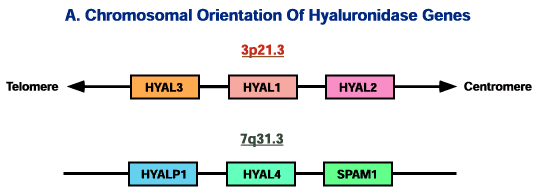

Fig. 2 6つのヒアルロニダーゼ遺伝子の2カ所のそれぞれの染色体の部位における配置図とそれらの遺伝子産物についての表。
第7染色体上の遺伝子の相対的な遺伝子のならびは確定されたが、セントロメアとテロメアへの方向は決定されていない。図は一定の尺度でもって描かれてはいない。
Clustal W プログラム13を用い、5個のヒアルロニダーゼ遺伝子cDNAの予測される翻訳産物とヒアルロニダーゼの偽遺伝子である、HYALP1の3個の「エキソン」の仮想的な翻訳産物を並べた(Fig. 3)。保存されたブロックはおそらくヒアルロン酸基質との結合と酵素活性に重要な領域を示すであろう。
Fig. 3
これまでに同定された5つのヒトヒアルロニダーゼすべてのcDNAの予測される翻訳産物と、ヒアルロニダーゼ偽遺伝子の3つの”エキソン”の仮想的な翻訳産物をCLUSTAL W プログラムを用いて作成したアラインメント。マウスHyal-1、Hyal-2、PH-20のシークエンスも示してある。同一のアミノ酸は四角で囲み、同族アミノ酸は影を入れた。保存されたブロックは、多分、酵素活性に重要であることを示すが、その領域が全体に見られる。
ハチ毒の酵素は363アミノ酸にわたってPH-20とHyal-1に約30%のアミノ酸の相同性を有する(Fig. 4)。この3種の酵素のアミノ酸配列にはアミノ末端にシグナル配列と開裂配列が共に存在する。PH-20には膜貫通型の脂質アンカーから酵素を遊離する開裂部位がもう一つ存在する。ハチ毒酵素はHyal-1ではEGFドメイン、PH-20では細胞接着部位と予測される脊椎動物の酵素のカルボキシ末端側の三分の一を欠く。PH-20のカルボキシ末端の膜貫通GPIアンカーはHyal-4にも存在する(図示していない)。Hyal-1のEGF様ドメインはショウジョウバエのserrateタンパク、アフリカツメガエルのXotchタンパクと相同性を有する。また軸索のガイダンスに関与するヒトのslit2タンパクに弱い相同性を有する。
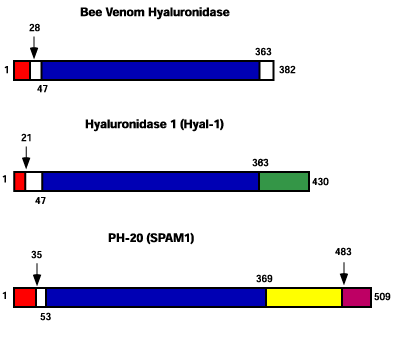
Fig. 4 ハチ毒ヒアルロニダーゼ、Hyal-1(血清ヒアルロニダーゼ)とPH-20(精子ヒアルロニダーゼ)の推定上のドメイン。
後者はまた、SPAM1(sperm adhesion molecule 1)としても知られている。3つの酵素の間でもっとも相同性の高い領域を青色で示す。ほ乳類酵素のカルボキシ末端の1/3は、相同性がほとんどない領域であるが、ハチ毒酵素にはない。透明帯結合ドメインはPH-20のカルボキシ側に存在する。PH-20と同様、EGF様ドメインがHyal-1のカルボキシ側に含まれる。この配列はショウジョウバエのserrateタンパク質(66アミノ酸にわたり52%)とアフリカツメガエルのXotchタンパク質(72アミノ酸にわたり35%)に相同性を有する。また、軸索のガイダンスに関与するヒトSlit2タンパク質(146アミノ酸にわたり34%)にも弱い相同性が見つかっている。相対的なポリペプチドの長さは一定の尺度で描かれている。
ハチ毒ヒアルロニダーゼの結晶構造解析の結果から、多くのグリコシダーゼによく見られるベータ/アルファ構造の繰り返しからなる円筒構造が示された。ハチ毒ヒアルロニダーゼタンパクの表面に観察される大きな溝は、ヒアルロン酸6糖をとどめるのに十分な大きさで、多くの芳香族ならびに疎水性アミノ酸残基を含み、おそらく、基質の結合と触媒作用に関与するものと思われる (Z. Marcovic-Housley私信)
対応するマウス遺伝子がマウス染色体の相同領域である9F1-F210と6A2(未発表)に見つかっている。ヒトとマウスの相同遺伝子同士の間の相同性は6つのヒトのパラロガス遺伝子間よりもはるかに高く、パラロガス遺伝子間の分岐は、ヒトとマウスが分岐する(約8000万年前)よりずっと以前に起こったと考えられる。これまで、ヒトとマウスで同定されたほ乳類ヒアルロニダーゼ遺伝子の系統的関係をFig.5に示す。樹状図は第3染色体上の3つの遺伝子は、第7染色体上の2つの遺伝子と1つの偽遺伝子よりはお互いがより緊密に関係していることを示している。3連の配列を生じた遺伝子重複とen masse重複が進化の中でいつ起こったのかを確立することも興味あることであろう。線虫C. elegansの遺伝子は脊椎動物のヒアルロニダーゼファミリーに相同性を持つ、ただ一つのヒアルロニダーゼ様配列を含む。興味深いことに、C. elegansに存在する遺伝子の2/3しか持たないショウジョウバエはヒアルロニダーゼ様配列を持たない。
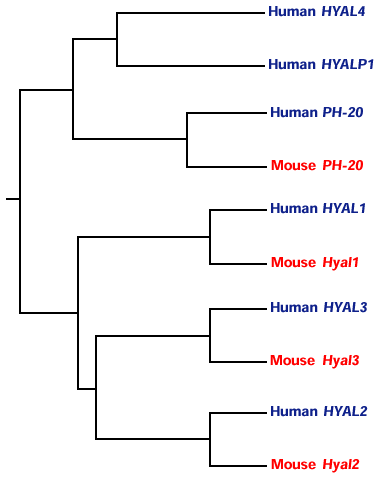
Fig. 5 6つのヒト、4つのマウスヒアルロニダーゼの系統解析。
樹状図はhttp://www.bionavigator.com.のBionavigator web siteで作成した。樹状図を作成するのに用いたプログラムはClustalW、dnadist、Neighbor、DrawGramである。
すべてのヒアルロニダーゼ遺伝子は特徴的な組織特異的発現パターンを持つ。偽遺伝子HYALP1が存在し、その中にある変異の特徴から、この多重遺伝子ファミリーはいまだ活発に進化しつつあることが示唆される6つのすべてのヒアルロニダーゼ配列の遺伝子構造をFig.6に示す。第3染色体上の遺伝子はすべてエキソン、イントロンに関して類似の構造を持つが、第7染色体上の遺伝子間ではほとんど保存されていない。さらに、第7染色体上の遺伝子は非コード配列の割合が高いので第3染色体上の遺伝子より有意に大きい。もっとも小さい遺伝子はHYAL1で3.7kbで、もっとも大きい遺伝子はHYAL4で32kb以上でほとんどその遺伝子サイズは10倍である。HYAL1は時々、そのmRNA中のエキソン1部分にイントロン1を含んでいる。それは他の遺伝子には見られない。

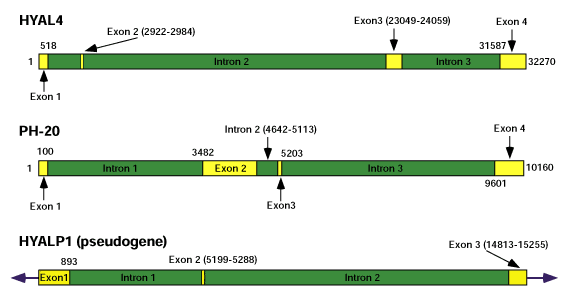
Fig. 6 ヒアルロニダーゼ遺伝子の遺伝子構造。
エキソンは黄色の長方形で示し、イントロンは暗緑色の長方形で示す。番号は他に示さない限りエキソンのヌクレオチドの開始と終止を示す。タンパク質に翻訳されないHYAL1mRNAアイソフォーム(本文参照)において、イントロン1がエキソン1の中に含まれている(明緑色の長方形で示す)。エキソンとイントロンは個々の遺伝子に関しては一定の尺度で示す。しかし遺伝子サイズは、その大きさにほぼ10倍の広がりがあるので、相対的な尺度で描かれていない。第3染色体上のヒアルロニダーゼクラスターの遺伝子はすべて似かよったエキソンとイントロンの構造をしているが、第7染色体上ではこの構造は保存されていない。
Hyal-1酵素の遺伝子は初めて同定された生体内のヒアルロニダーゼであった。もう一つの広く発現している酸性条件下で活性を持つヒトヒアルロニダーゼ、Hyal-2は染色体上で隣接する遺伝子にコードされている14。Hyal-2は珍しい基質特異性を持ち、高分子量のヒアルロン酸を約20kDaの中間のサイズの断片に分解する。多分、この2種のヒアルロニダーゼは共同して働き、Hyal-2消化の生成物がさらに低分子量のオリゴ糖にHyal-1の働きによって分解されるのであろう。3p21.3の遺伝子座にコードされる第3の酵素であるHyal-3についてはほとんどわかっていない。ほ乳類の精巣と骨髄に強い発現がハイブリダイゼーションによりみられる。この二つの組織は動物が生きている間、幹細胞様の段階を維持していることから、Hyal-3ははじめ胎児期に発現されるヒアルロニダーゼであることが示唆される。精子に存在する酵素であるPH-20とHyal-3は精巣で高い発現を示すが、Hyal-3が受精の過程に関与するかどうかは不明である。
PH-20タンパク質は単クローン抗体によって精子の先体膜上で初めて同定された。ハチ毒ヒアルロニダーゼとの相同性から、卵細胞を包むヒアルロン酸に富む卵丘への侵入と受精の両方に必須なヒアルロニダーゼとして同定された。PH-20はGPIアンカーによって膜に結合した大きなポリペプチド鎖として合成される。それから、プロセッシングを受け、ジスルフィド結合によって結合した2本のポリペプチド鎖の可溶性タンパク質となる。卵丘への結合と侵入、ヒアルロン酸の分解と受精、の過程における二つの精巣ヒアルロニダーゼ酵素の相対的な寄与がどのようなものかについては不明である。しかしながら、1)ヒト尿中にPH-20の2つのフォームに匹敵する2つのフォームのHyal-1がみつかったこと、2)親分子もプロセッシングを受けた分子も酵素学的には活性があることなどは興味深い。キチナーゼのヒトの類似分子がマクロファージに見つかり、それはヒアルロニダーゼのようにエンド-β-N-アセチルグルコサミノダーゼで、同じような方法でプロセッシングを受ける。これらすべての酵素の2種のフォームの意味に関しては不明である。両者は互いに匹敵する比活性を有するので、典型的な酵素源と活性酵素の関係ではない。
新たなヒアルロニダーゼのパラログであるHYAL4は 7q31.3染色体に同定された。HYAL4 cDNAは2414塩基対の長さであると算出された。予備的な証拠ではHyal-4はコンドロイチナーゼであって、ヒアルロン酸に対しては活性を持たない(未発表)。これは脊椎動物組織で初めての酵素であるコンドロイチナーゼであると同定された。この遺伝子ファミリーにコンドロイチナーゼ活性がコードされることは、脊椎動物のヒアルロニダーゼが、その活性は低いがコンドロイチン硫酸を分解する活性を有する事を考えると、驚くに当たらない。コンドロイチン硫酸はヒアルロン酸の近縁の分子であり、違いはN-アセチルグルコサミンがN-アセチルガラクトサミンで置き換わっているところである。両ポリマーはすべてβ結合である。コンドロイチン硫酸には硫酸化があるが、このランダムに硫酸化されたポリマーの硫酸化を受けていない、または、硫酸化が低い部位が実際の開裂部位である可能性がある。
3番目の遺伝子であるHYALP1は2カ所の未熟な終結部位を生じるいくつかの欠失があることから偽遺伝子であることが判明した。ヒトの配列にわずかな変異しか見られないということからHYALP1はほんの最近偽遺伝子に退化し、他の種では機能的であるかもしれない。HYALP1のエキソンの仮想的翻訳産物を他のヒアルロニダーゼとの相同性を示しながらFig.6に示してある。エキソン1の変異により欠失したアミノ酸とエキソン3の未熟な終結部位はXで示した。
ヒアルロン酸の代謝は非常に活発である。ヒアルロン酸レベルの急激な上昇は、じんま疹、創傷治癒と炎症を伴う浮腫、移植後に起こる臓器肥大など、多くの臨床場面で起こる。梗塞の後に起こる心筋の傷害は、部分的には外傷後の組織膨張に伴う圧力が関与した壊死による。このようなショック、敗血症、火傷の患者では、血中ヒアルロン酸のレベルが急速に上昇する。このようなヒアルロン酸レベルの上昇は、その大きな保水力により、血液量を増加させ、循環系がつぶれてしまうのを遅らせるという、ほ乳動物の生存メカニズムの一つなのかもしれない。ヒアルロン酸とその結合水は、完全に独立したコンパートメントを形成していて、純粋に体積増加機能をもった他のコンパートメントとは平衡状態にはないかもしれない。このような状況でのヒアルロン酸レベルの上昇はストレス反応の一つである。ヒアルロン酸の早い代謝レベルを中断すること、それは多分ヒアルロニダーゼ阻害剤を介するメカニズムであろうが、それは引き続きストレス応答性サイトカインによって調節され、この機構はこのような極度の臨床場面で、ヒアルロン酸レベルを即時に上昇させるための迅速な応答機序をほ乳類に提供した。予備的な証拠からこのようなシナリオが得られている (MioとStern、未発表)。ヒアルロニダーゼに分類される酵素とその阻害剤は、前述または他の臨床場面に介入する主要な治療的価値があるかもしれない。
異なったヒアルロニダーゼの遺伝欠損は臨床的に異なった症候群をひきおこす可能性がある。事実、ヒト血清ヒアルロニダーゼであるHyal-1の場合はすでにムコポリサッカリドーシスIXと名付けられ報告されている。Hyal-1活性の消失で、この患者に観察されるように臨床的特徴が穏和であることは、ヘモグロビン異常症のいくつかでみられる類似物の場合のように、胎児ヒアルロニダーゼであるHyal-3の持続によって代償されているのかもしれない。"Michelin tire"症候群の新生児の場合、逆位が見つかっている7qの領域の切断部位の近傍にHYAL4は存在し、この疾患の候補になっている。
ヒアルロニダーゼは、また、癌が浸潤し、転移拡散していく機構としてももたらされる。癌細胞を取り巻くヒアルロン酸のレベルは、しばしば、癌の悪性度と予後の悪さに相関する。ヒアルロン酸の過剰な合成はanchorage-independentな増殖を促進する。ヒアルロニダーゼ活性の消失は多段階な腫瘍発生過程に必要とされるいくつかのステップの一つである。HYAL1遺伝子(また、LUng CAncer-1、LUCA-1として知られている)はhomozygous deletionと機能的な癌抑制活性によって推定された候補としての癌抑制遺伝子座にマップされる。この部位の半接合体または二つの対立遺伝子の一方のこの部位の欠失が、多くの口腔、頭部、頸部、肺の癌にみられる。残りの対立遺伝子の変異はホモ接合体を生じるために必要であろう(両対立遺伝子における機能消失)。酵素にとって、本来触媒作用を持つものであるから、二つの対立遺伝子のうちただ一方を失っただけならば有害ではなく、残りの対立遺伝子が細胞に十分なレベルの活性を供給しうると推測される。影響を及ぼすには両対立遺伝子の欠損すなわちホモ接合体の欠損が必要であろう。しかしながら、HYAL1遺伝子の変異は広範な調査にも関わらず癌には見つかっていない。癌抑制遺伝子(TSG)の古典的な概念はこのTSG候補領域には適用できない。
しかしながら、我々は最近、HYAL1遺伝子産物を機能的に不活化させる領域を同定した。多くのがん細胞株でヒアルロニダーゼ活性、または、Hyal-1タンパク質は検出されなかったが、Hyal-1のmRNAは存在していた。通常見られないほど大きな5'UTRは保持されたままのイントロンがあることを示している(Fig.6)。保持されたイントロンは正しい開始メチオニンコドンにリボソームが結合するのを阻害することにより翻訳を停止させた。このような癌における遺伝子の不活化の様式がどれほど広く行き渡っているかはわかっていない。しかしながら、この発見から癌の発達の過程でのヒアルロニダーゼの消失の重要性が強調される。ヒアルロニダーゼの消失は癌細胞にヒアルロン酸の豊富な環境を提供し、それが増殖、移動、そして転移拡散を促進する。これらは悪性度を獲得する好条件となる。癌細胞は、多分、望ましくない遺伝子産物を黙り込ませてしまうように広いレンジの機構を適応させたのであろう。明らかに、癌抑制遺伝子の不活性化というイベントは遺伝子DNAのレベルだけでなく、HYAL1の場合のようにmRNAのレベルでも起こりうる。7q31.3遺伝子もまたTSG領域にマップされることは注目されるべきである。
ヒアルロン酸の代謝は非常に速いが、その代謝のメカニズムの詳細のほとんどは分かっていない。ほ乳類の循環系の中でなぜヒアルロニダーゼのみが酸性領域活性酵素なのかわかっていない。Hyal-1は組織で合成され、他の組織に取り込まれるために血流で輸送されるのであろうか?酸活性ヒアルロニダーゼは本当にリソゾーム酵素なのだろうか?もしそうなら、このようなヒアルロニダーゼの至適pHが、リソゾームのpH4.5より有意に低いpH3.7なのはなぜだろうか?ムコリピドーシスIIとIIIの患者の証拠が示すように、彼らのHyal-1は普通ではなく、また大抵のリソゾーム酵素とも異なっている18。これらの遺伝疾患を持つ患者血清中ではその活性は上昇していないことから、Hyal-1はリソゾームへターゲットされていないか、通常のリン酸化マンノース残基を認識する経路以外の機構によってターゲットされていることが示されている。培養細胞では、ストローマ細胞であれ上皮細胞であれほとんどのヒアルロニダーゼ活性は迅速に培養上清に分泌され細胞層にはとどまらない。これは生体内の状況を反映するのであろうか?ヒアルロニダーゼは分泌され、そしてそれから二次的に細胞に取り込まれリソゾームへ届けられるのであろうか?このような状況は培養細胞によって試験管内で再現されているとはいえない、なぜなら酵素は直ちに大量の培養液によって希釈されてしまうからである。
受容体を介したエンドサイトーシスが細胞によるヒアルロニダーゼの取り込みに関与するのであろうか?もし、そうならこれらの受容体の実体とはなにであろうか?CD44はヒアルロン酸の取り込みと分解に関連づけられてきた。どのようにしてCD44はヒアルロニダーゼによるヒアルロン酸の取り込みと分解の全課程に関与するのであろうか?多分その多様なバリアントエキソンフォームの一つであろうが、CD44はヒアルロニダーゼまたはヒアルロン酸−ヒアルロニダーゼ複合体の受容体なのであろうか?このような機構を考える時の難しさは酸活性ヒアルロニダーゼは中性ではヒアルロン酸基質に結合しないことである。もしかすると、中性で酵素−基質結合を促進するのに関与するもう一つの分子または補因子があるのであろうか?
皮膚は全生体内ヒアルロン酸量の50%を含むが、ヒアルロン酸の半減期は約一日で、軟骨のように見かけ上不活性な組織でさえ、ヒアルロン酸代謝の半減期は1-3週間である。血流中ではヒアルロン酸の半減期は2-5分である。このような異化作用のすべては多分ヒアルロニダーゼの結果であると考えられている。このように非常に異なった代謝速度を統合している調節機構はなんなのであろうか?脊椎動物のヒアルロン酸は分子量もいろいろで、細胞外にあったり、自由に循環していたり、細胞または組織に緩く会合していたり、軟骨のようにプロテオグリカンが豊富なマトリックスに強固に織り込まれていたり、受容体によって細胞表面に結合していたり、いくつかの細胞内局在を示すなど、多くの状態をとりうる。これにさらに付け加えられているものはヒアルロン酸結合タンパク質、または、hyaladherinsである。これらのように物理的、化学的に広範にわたる状態のヒアルロン酸基質に対してその異化機構はどれほど異なるのであろうか?ヒアルロニダーゼ活性は生体内では細胞外マトリックス(ECM)に活性型でとどまることはないようである。もしそうなら、重大な破壊をひき起こすに違いない。ECMに見つかるとすれば、それは不活性型か、多分、インヒビターに結合した形の抑制型であろう。このような状況はメタロプロテアーゼ(MMP)とMMP活性を精密に制御しているTIMPとの関係に類似しているかもしれない。
ヒアルロニダーゼの発現レベルの調節に関しては、わかりはじめたばかりである。培養細胞では、ストローマでも上皮細胞でも、ヒアルロン酸とヒアルロニダーゼは逆のパターンを示す。培養皿にまばらにまいた細胞は増殖し、培養皿を埋め尽くすようになる時には、ヒアルロン酸レベルは高く、酸活性ヒアルロニダーゼ活性はほとんど検出されない。細胞が接触阻害を起こし、増殖を停止するようになると、ヒアルロニダーゼ活性は急激に上昇し、ヒアルロン酸蓄積のレベルが急速に低下する19。このような情況は形質転換した細胞では見られない。ヒト皮膚より得られたケラチノサイトは培養下でカルシウムレベルを0.05から1.30mMに上昇させると分化させることができる。高カルシウム培地は細胞を層状化させ、ケラチンを合成するようにする。分化に伴い、ヒアルロン酸の合成と蓄積のレベルは低下し、 Hyal-1ヒアルロニダーゼの合成は25倍にまで上昇する6。明らかになってきた図式としては、未分化な速い速度で増殖する細胞は高いヒアルロン酸レベルを有するが、ヒアルロン酸リッチマトリックスを失った細胞は、細胞増殖が遅くなり、分化のプログラムを開始するというものである。ヒアルロニダーゼ活性はこのような遷移過程を仲介する鍵なのかもしれない。
近年、真皮線維芽細胞がマイクロマス培養系で軟骨細胞に分化する過程でのヒアルロニダーゼレベルが調べられた22。われわれは分化に伴うHyal-1の劇的な増加、Hyal-2、Hyal-3の中程度の増加を見いだした。いろいろな単離された臓器や組織の系での分化の過程は、はじめにBryan Tooleによって発生学におけるヒアルロン酸の役割の彼の古典的な研究で提案されたように、ヒアルロニダーゼの調節を行う多様な機構を研究する多くの機会を提供しうる(このシリーズのB. Tooleによる総説を参照)。
ヒアルロン酸異化作用の領域には多くの他の未解決の疑問が存在する。なぜ二つのフォームのHyal-1酵素があるのか?エンド型蛋白分解酵素反応を引き起こすプロテアーゼの実体はなんなのか、そしてこのような開裂反応はどこで起こるのか?Hyal-1の二つのフォームは膜結合型と可溶性型のPH-20の二つのフォームに類似しているのか?Hyal-1の二つのフォームのうちどちらがin vivoでより活性か?培養細胞は両者を発現しうるが、尿中には両者が見いだされるのに血清には一方だけなのはなぜか?尿中の非常に高いヒアルロニダーゼ活性、それはすべてHyal-1のようであるが、その意義は?ほ乳類の生体内で毎日分解されるグラム単位のヒアルロン酸を説明するようなまだ発見されていない他のヒアルロニダーゼがあるのか?70kgのヒトは15gのヒアルロン酸を所有し、毎日1/3が代謝される。ヒアルロン酸代謝は明らかに我々がほとんど知らない時間的、空間的メカニズムによって厳密にコントロールされている。多分、この6つからなるファミリーに無関係で、in vitroの酵素アッセイ固有のアーティファクトのために今日まで同定されていない他のヒアルロニダーゼは存在するであろう。明らかにほ乳類ヒアルロニダーゼについては多くの問題が残されている。



