
氏名:Scott Strong
Scott Strongは、Cleveland Clinic Foundation (Cleveland、オハイオ州)の結腸・直腸の外科医であり、Lerner研究所の免疫部門と共同の役職にある。彼はアイオワ州立大学で化学工学のB.S.を受け、アイオワ大学からM.D.を受けた。その後、1990年にCleveland Clinicで結腸・直腸の外科医の研修を始める前、ミシガン州立大学のButterworth病院で外科のレジデントを終了した。加えて、彼は炎症性腸疾患の研究の分野の第1人者であるClaudio Fiocchi博士と共同研究を行った。1993年に現在のポストに就いた時、彼は炎症性腸疾患の病因の細胞メカニズムを調べることに専念するための研究室を作った。彼は腸粘膜における免疫細胞・非免疫細胞の相互作用の重要性を強調して、最近では新しいウイルスに誘導されたヒアルロン酸とCD44を介した細胞間相互作用のメカニズムに焦点を絞った研究をしている。

氏名:Carol de la Motte
Carol de la Motteはコーネル大学、(Ithaca、ニューヨーク州)で微生物学専攻でB.S.を受けた。彼女はレキシントンのケンタッキー大学で医学真菌学を専攻して医学微生物学の大学院を卒業した。1978年に彼女はCleveland Clinicの研究部門に入り、彼女が非免疫細胞と免疫細胞の相互作用に影響する因子に興味を持ったのがその後の研究の始まりである。Max Proffitt博士の研究室で彼女はウイルス-リンパ球の相互作用を調べ、これが免疫細胞が間葉性細胞を殺すのにどのように影響するかを調べた。次に、彼女はPaul DiCorleto博士の研究室に移り、そこで内皮細胞に発現するリンパ球接着分子とそれらの動脈硬化と炎症性疾患における意義に興味をもって研究した。数年前、彼女は炎症性腸疾患における間葉細胞の関与の細胞メカニズムを調べるために結腸・直腸外科部のScott Strong博士の研究に参加した。思いもかけず、この研究は彼らをヒアルロン酸へと導き、Vincent Hascall博士のチームと大変面白く実り多い共同研究へと導いた。
恒常性とは化学成分のバランスによって維持される生理的な平衡状態を指し、秩序だった臓器系において機能する。それぞれの臓器の固有な働きは、様々な発生起源の無数の細胞が協調して働くことを必要とする。高等生物の恒常性を維持するように編成された細胞の活動は、その数と複雑さにおいて驚くべきものがある。本シリーズToole論文では、ヒアルロン酸の組織機能維持における役割のみならず、諸臓器の形態形成における不可欠な役割が述べられている。
組織内の細胞集団は環境内の変化(例えば、物理的障害、化学物質の摂取や吸入、温度の変化、細菌感染)に対してうまく対応できるように組織化されており、必要に応じて修復し、組織を維持することによって生体を守っている。生体全体のみならず個々の臓器の必要性に対して、細胞は動的且つ相互にコミュニケートしていなければならない。
免疫系のリンパ球と神経系の神経細胞は、異なった細胞集団とコミュニケートし相互作用するよく知られた細胞の例である。しかしながら、細胞が静的で単一の働きしかしていないという既成概念はもはや通用しない。内皮細胞はかって血管を単純に裏うちしているだけと考えられていたが、周辺の細胞とコミュニケートできるような化学シグナル物質(サイトカインや成長因子のような)を放出したり、応答したりすることがわかっている。加えて、内皮細胞は、リンパ球接着分子を細胞表面に発現し、循環血球細胞との物理的な相互作用を促進している。これらの細胞接着分子は、下方組織中へのリンパ球の浸潤を完全に規制しており、局所炎症反応のコントロールに関与している。最近になって、繊維芽細胞や平滑筋細胞のような組織を構築する細胞も白血球接着分子が介する免疫系細胞との相互作用をするばかりでなく、化学シグナル物質を同様にやりとりすることが証明された。
ヒアルロン酸も環境変化が引き起こす細胞の応答に関与している。本シリーズの論文でもa、最終的に再狭窄に至る物理的血管障害に平滑筋細胞が応答する時のヒアルロン酸の役割が述べられている。ヒアルロン酸はまた微生物の侵入に対する細胞の反応にも関与しうる。平滑筋細胞のウイルス感染は、例えば感染していない細胞にはみられないヒアルロン酸の応答を起こさせる。このようなヒアルロン酸の応答(下記のような)が平滑筋細胞の白血球との相互作用を助長して、炎症に寄与している1。
a このシリーズのWightの総説を参照せよ
大腸は食物が腸管を通る時、消化物の不要部分から水分を吸収することを主機能とする臓器である。ヒアルロン酸は結腸組織に中等量に存在し、組織浸透圧調節に重要な機能を果たしている。結腸は弾性のある管であり、その内部にある粘膜層の組織は血管の組織といろんな点でよく類似している(Fig. 1A)。結腸の内腔側表面は単層の上皮細胞で被われており、その下に粘膜固有層というコラーゲン線維と平滑筋細胞の束よりなるネットワークがある。このネットワークのすぐ下に粘膜筋層という平滑筋細胞の薄い層がある。この内側の粘膜層の外側にはさらにマトリクスと筋固有層という厚い筋肉細胞層がある。 通常は、結腸粘膜は白血球の細胞集団としてT細胞とB細胞、形質細胞、組織球、肥満細胞を含んでおり、これらの白血球は粘膜固有層に規則的に分布する毛細管を通って到達する。これらのリンパ球は粘膜を監視し、結腸の内腔側にある異物に対する免疫防御をしている。粘膜リンパ球は、おそらく粘膜筋層の近くにあるリンパ管を経て血流に再び入り、特異的抗原刺激によって問題の生じた領域へ呼び戻されるまで血液リンパ系の臓器を自由に循環している。
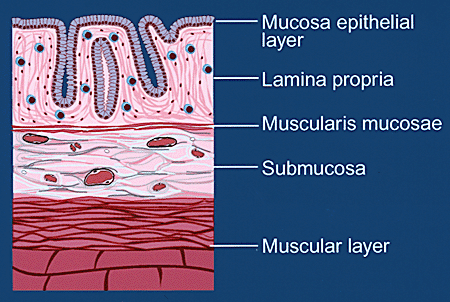
Fig. 1A 結腸の組織層の模式図 (A)正常
結腸癌と炎症性腸疾患は大腸の病気として比較的よくみられる二大重篤疾患である。結腸ガンは老人によくみられる悪性の疾患であり、他の臓器によく転移する。逆に、炎症性腸疾患は若年者の良性疾患であり、腸のみならず腸以外の部位に炎症が起こるのが特徴である。ある種の結腸癌が転移できるかどうかはヒアルロン酸レセプターCD44の発現と関連しており、これはヒアルロン酸と癌の進行の関係を示唆する。CD44とそのいくつかの亜型がしばしば多くの結腸癌に過剰発現しているb。亜型の一つであるCD44v6は一般に腫瘍の進行した段階で発現して、結腸・直腸癌に関連した死亡と強く関係している2。最近、腸の間質細胞が腺癌が産生する因子に応答してヒアルロン酸を合成することが報告されている。
b このシリーズのKnudsonの総説を参照せよ
炎症性大腸疾患の原因は外因子的であるようにみえるが、環境因子や微生物学的因子が遺伝的可能性のあるヒトの腸の免疫応答を引き出すように思われる。ヒアルロン酸がこの炎症性の疾患において中心的な役割をしているらしいといういくつかの証拠がある。ヒアルロン酸が増加することが炎症性腸疾患の患者の腸で見られている。他の炎症性疾患と比較して、CD44の増加が炎症性腸疾患の一つである潰瘍性大腸炎の患者の炎症を起こした結腸組織に広範に認められ、とりわけ上皮細胞にCD44v6とCD44v3の発現が増大している。興味あることに、炎症性腸疾患の腸管以外の組織では30から50%の患者で見られ、関節、皮膚といったヒアルロン酸が多い組織で同じように炎症が起こっている。
炎症性腸疾患の病因に関連した免疫現象が良く調べられており、それらの所見は健常者と患者の応答に違いがあることをはっきり示している。例えば、特異的サイトカインを産生するリンパ球の分布が健常者とクローン病や潰瘍性大腸炎の患者で異なっていることはよく知られている。しかしながら、微生物がこれらの病気の進展にどのように影響するのかはほとんど知られていない。ウイルスが炎症性腸疾患の病因になっているかもしれないという推論は、気道ウイルスの感染が引き続き病気の憎悪を伴うという臨床所見と一致するためかなり長い間提唱されてきた。
炎症性腸疾患により粘膜の免疫細胞集団は劇的に増殖し、浸潤細胞の大部分が単核白血球で構成されるようになる(Fig. 1B)。さらに、平滑筋細胞の過形成と細胞外マトリクスの蓄積のため粘膜筋層は正常の約300倍近くの厚さにもなる。このような特徴は集合した白血球と間葉系平滑筋細胞が炎症性腸疾患の進行と広がりにおいて重要であるということを示唆している。
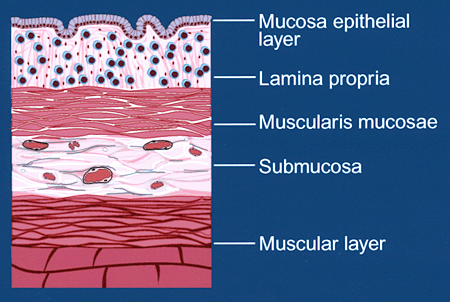
Fig. 1B 結腸の組織層の模式図 (B)炎症性腸疾患
白血球の産生する炎症性サイトカイン、例えば腫瘍壊死因子(TNF-α)とインターロイキン1は結腸平滑筋細胞の増殖を引き起こし過形成に寄与する。 加えて、TNF-αを代表とするこれらのサイトカインは血管接着分子-1(VCAM-1)として知られている白血球接着分子の発現を平滑筋細胞に増加させ、このタンパクによって単核白血球を平滑筋細胞の表面に接着させる(Fig.2)。ある種のリンパ球のサブセットがその接着レセプターVLA-4(晩期抗原-4)を介してVCAM-1に接着する作用は、これらのリンパ球の72-kDのゼラチナーゼの産生と活性化を引き起こし、組織の再構築の役割を果たす。
興味あることに、ウイルスは結腸平滑筋細胞を含むさまざまな細胞のVCAM-1を増加させる。しかしながら、ウイルスが引き起こす白血球と平滑筋の相互作用の機作は炎症性サイトカインによるそれとまったく異なっている(Fig.2)。
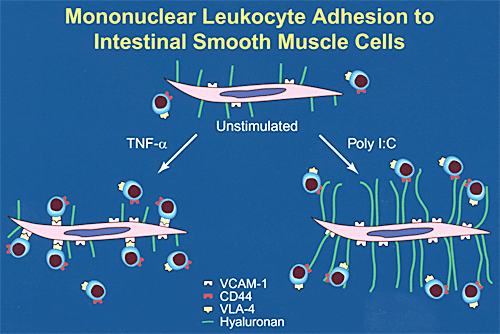
Fig. 2 炎症性サイトカイン(TNF-a)とウイルス(ポリI:C)によって誘導された結腸平滑筋細胞表面上の白血球接着分子への白血球接着の模式図
培養ヒト結腸平滑筋細胞は気道腫瘍ウイルス(RSV)と麻疹ウイルスに高い感染性があり、これがこれらのウイルスが炎症性腸疾患に関与していることの状況証拠となる。平滑筋細胞は正常な機能の一部として本来ヒアルロン酸を合成しているがウイルス感染に反応して単核白血球(すなわちT細胞、B細胞、単核細胞、肥満細胞)に対してきわめて接着しやすいヒアルロン酸を細胞表面に産生する。主要なヒアルロン酸受容体であるCD44は、この細胞間相互作用に最も大きく関与する白血球リガンドである。
位相差顕微鏡で観察すると、RSV感染後一日で結腸平滑筋に白血球の接着に劇的変化が見られる(Fig.3)。白血球は平滑筋細胞の灰色層に接着した明かるい球体として見える。ウイルス感染を模した合成二重ラセンRNAであるポリI:Cは、細胞を殺すことなく沢山のウイルス応答経路を活性化するが、これは結腸平滑筋に対して対してのみならず、同様に血管(腸間膜動脈)や気道(気管支)組織から単離した平滑筋細胞に対しても白血球の接着を誘導する(Fig.4)。
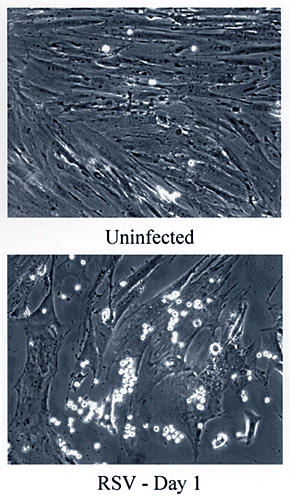
Fig. 3 気道腫瘍ウイルス(RSV)感染による結腸平滑筋細胞への白血球の接着。
コンフルエントの結腸平滑筋細胞をRSV含有、非含有培養液で37℃で18時間処理した。非刺激白血球を1時間接着させた。非結合白血球を洗い去り、位相差顕微鏡で観察(100倍)。白血球は培養プレートに接着した平滑筋細胞の上に鮮明な球として見える。
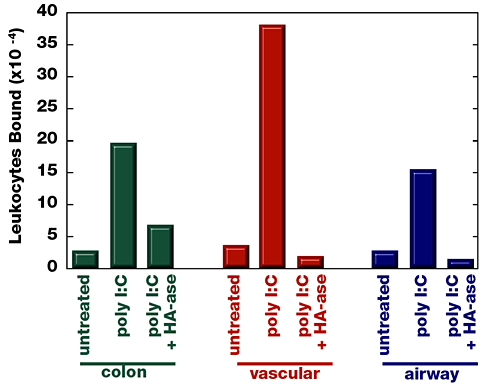
Fig. 4
ポリI:C処理は結腸、血管、気道の平滑筋細胞への白血球の接着を増加させ、これはヒアルロニダーゼ消化で消滅した。コンフルエントの平滑筋細胞をウイルス擬似物(ポリI:C)を含む、または含まない培養液で一晩処理した。精巣ヒアルロニダーゼを定められた培養皿加え、37℃で10分培養。放射性標識された白血球の結合を示す。
大部分の白血球の接着にはウイルスあるいはポリI:C処理平滑筋細胞表面上のヒアルロン酸が介在している。Fig.4に示すように、ヒアルロニダーゼ処理により大部分の白血球の接着は阻害される。残りの白血球接着は平滑筋細胞上のVCAM-1に媒介されており、上記のように特異的な抗VCAM-1モノクロナル抗体により阻害される。患者間の変動は全単核白血球接着に対するそれぞれの接着分子の相対的関与として説明できる。
上記のように、ウイルス感染やポリI:C処理はVCAM-1(通常はTNF-aによって誘導されるよりも高濃度に)とヒアルロン酸を平滑筋細胞表面の発現を増加させるが、ヒアルロン酸のみが白血球の接着を有意に媒介する。細胞表面からヒアルロン酸を酵素で除去すると、VCAM-1が十分に白血球に結合できるようになる。これらの知見はヒアルロン酸のもう一つのの興味ある機能、すなわち他の接着分子を覆いかくすという機能を示唆している(Fig.2)。培養平滑筋細胞に対するヒアルロン酸媒介による白血球の接着は、VCAM-1による白血球の接着とは明らかに異なっているようにみえる。位相差顕微鏡による観察ではポリI:C処理の平滑筋細胞がヒアルロン酸を介して白血球に結合していることを明示している(Fig.5)。細胞数個の距離をへだてた白血球の鎖状の形成が認められる。ヒアルロニダーゼによりヒアルロン酸を除くと、典型的なVCAM-1による単独の白血球の接着像があらわれる。

Fig. 5 ポリI:C誘導白血球の結腸平滑筋細胞への接着の位相差顕微鏡像。
ポリI:C処理した平滑筋細胞への白血球の接着は、しばしば鎖状あるいは塊状で並んでいる。白血球接着前のヒアルロニダーゼ処理はもう一つの分子VCAM-1を露出せしめ、右端のパネルに示した明らかに異なった接着パターンとなった。
培養結腸平滑筋細胞は、通常の条件下では合成しているヒアルロン酸のわずかな量(15時間培養で20%ぐらい)を保持するに過ぎない。ポリI:C処理に応答して、ヒアルロン酸の全合成が高まり、細胞の保持する比率も増加する(約35%ぐらい)。ポリI:C処理平滑筋細胞により合成されたヒアルロン酸は未処理の細胞ではみられない2つの異なった構造上の型に組み込まれる。共焦点顕微鏡で結腸平滑筋細胞をヒアルロン酸(フルオレセイン)、CD44(テキサスレッド)で共染色して見ると、ポリI:Cは細胞数個の長さの範囲を持つ鎖状のヒアルロン酸構造のみならず細胞周囲のヒアルロン酸外被の産生を誘導しているのがわかる(Fig.6A)。平滑筋細胞はその表面に多量のCD44も発現させるが、この発現はポリI:Cによっては変わらない。
非活性の白血球(核が青に染色されることで同定できる)はその細胞表面にCD44(テキサスレッドの染色で同定できる)を同様に発現しているが、未処理の平滑筋細胞のヒアルロン酸には結合しない。しかしながら、非活性の白血球はCD44を通じて平滑筋細胞のウイルス誘導のヒアルロン酸によく結合し(Fig.6B)、また、この相互作用から白血球のCD44は撚糸状のヒアルロン酸構造にのみ結合するが細胞周辺のヒアルロン酸外被には結合しないという選択性があるようにみえる。
非活性の白血球の結合に適した巨大な分子構造へのヒアルロン酸の組み込みに関与する因子はまだ明らかにはなっていないが、いくつかのヒアルロン酸結合タンパク質が含まれるようである。可能性のある候補物質として血清タンパク質のインター-α-トリプシンインヒビター(I-α-IあるいはITI)があり、これは平滑筋細胞を含むさまざまな細胞のヒアルロン酸細胞周囲外被の安定化の作用があるといわれている。同様に、アグリカン、バーシカン、ヒアロネクチン、あるいはTNF刺激遺伝子-6(TSG-6)により産生されるタンパクなどのヒアルアドヘリン分子はヒアルロン酸を直鎖状構造に整列させるのに役立っており、おそらく白血球のCD44と相互作用している。
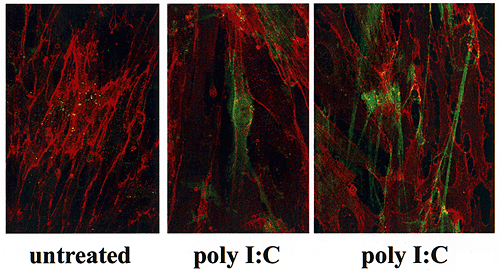
Fig. 6A 結腸平滑筋細胞のポリI:C誘導ヒアルロン酸とCD44の共焦点顕微鏡像。
結腸平滑筋細胞はヒアルロン酸とCD44に対して二重染色された。ヒアルロン酸結合タンパク質は二次蛍光標識体と共に用いヒアルロン酸は緑で示している。CD44を検出するために、CD44に対するモノクロナル抗体(A3D8)がテキサスレッド標識の2次抗体と一緒に使われた。
(A) 未処理の平滑筋細胞は細胞表面にほとんどヒアルロン酸(緑色)を持たない。
ポリI:C処理後に、繭様の細胞周囲のヒアルロン酸マトリクス(中図)が培養ではっきり見え、またいくつかの細胞におよぶ撚糸状のヒアルロン酸構造(右図)も見える。
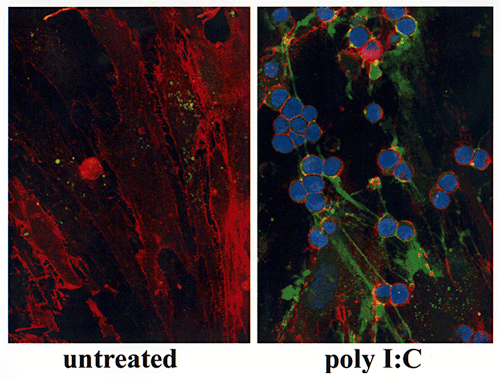
Fig. 6B
(B) 白血球(青の核染色)接着は撚糸状の構造で媒介される。白血球表面のCD44(赤色)はヒアルロン酸への接着を媒介する。
ヒアルロン酸をリガンドとして結合する主なレセプターであるCD44については多くのことが知られているb。最近の知見では炎症過程でのCD44レセプター/ヒアルロン酸の相互作用の潜在的重要性が強調されている。例えば、CD44のヒアルロン酸への特異的な結合は末梢血T細胞を活性化し、IL-2レベルを高め、単球細胞を活性化して炎症性サイトカインのIL-1やTNF-αを多量に放出させる。同様に、T細胞上のCD44の活性化は炎症部位への白血球の血管外脱出のために必要であり4、CD44は中枢神経系や実験的脳脊髄炎における炎症で観察される二次的白血球の集合にも重要である5。他の組織において、ヒアルロン酸のマクロファージへの結合はIL-12の産生のみならず6、ケモカイン(すなわち、RANTES、MIP-1a、MIP-1b)の発現を高めるように調節することがわかっている。腸平滑筋細胞においてウイルス介在で誘導されたヒアルロン酸は、CD44を介して非活性の白血球に結合するので、これに伴って白血球が活性化し、炎症反応を悪化させるようである。
ウイルスに感染した結腸平滑筋細胞が、非活性のリンパ球に対して接着性のある撚糸状ヒアルロン酸を形成する理由はほとんどわかっていない。最近の文献によると、CD44の活性化にはそれがヒアルロン酸に結合することに必須であり、しかしその必要性はウイルス感染で誘導される接着のメカニズムには不必要らしいことを示唆している。従来の白血球接着分子(例えば、VCAM-1)への結合とは異なり、撚糸状ヒアルロン酸構造への白血球の結合は、様々な種類の単核白血球がお互いに相互作用し、さらに免疫応答を可能にしている。ヒアルロン酸は近接して結合した白血球が直接細胞表面の接着を介して、あるいは間接的に可溶性サイトカインを産生を介してコミュニケートできるような微小環境を創り出しているのかもしれない。ウイルス感染で見られた新しいヒアルロン酸の応答は、腸疾患と慢性炎症疾患の病因をよりよく理解するためのさらなる調査研究にとって拍車をかけることになるであろう。



