|
|
αマンノシダーゼとEDEMタンパク質:糖タンパク質の小胞体関連分解に果たす役割 |
||||||||||||||||||||
|
糖タンパク質の小胞体関連分解 |
||||||||||||||||||||
|
||||||||||||||||||||
|
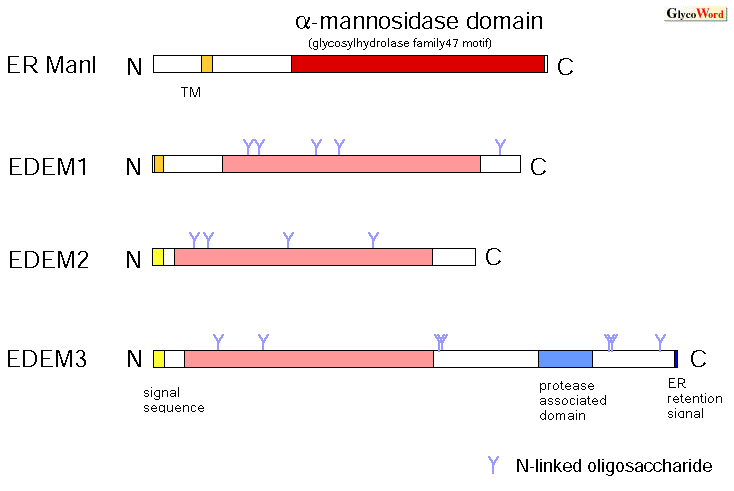
哺乳類のEDEM(ER degradation enhancing α-mannosidase-like protein)と酵母ホモログであるHtm1p/Mnl1pは、いずれもαマンノシダーゼ様のドメインをもつが酵素活性は検出されず、糖タンパク質のERADを促進する機能をもったタンパク質としてクローニングされた。その後データベースを検索することによって、哺乳類には3つのEDEMホモログタンパク質が存在することが明らかになった(1,2,3)。図2にER ManIとEDEMのドメイン構造を示す。いずれもクラスI α-1,2マンノシダーゼ (glycosylhydrolase family 47)モチーフをもっている。3つのEDEMタンパク質の間では、この領域において、それぞれ約45%という高いアミノ酸相同性を示すのに対し、ER ManIとEDEMタンパク質の間では約33%とやや相同性が低くなる。クラスI α -1,2マンノシダーゼファミリーにはもうひとつ、ゴルジマンノシダーゼI(Golgi mannosidase I: Golgi ManI)が一群のサブファミリーを形成しており、図3に示すようにそれぞれ、ER ManIとは約38%、EDEMとは約30%のアミノ酸相同性を示す(文献1の系統樹参照)。また、EDEM1,2,3に関して見てみると、EDEM3においてC末端領域がやや長く、Protease associated domainと呼ばれるモチーフが存在しているが、その機能は今のところわかっていない。 |
||||||||||||||||||||
|
||||||||||||||||||||
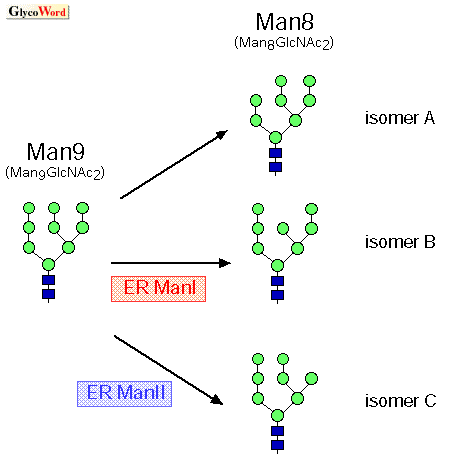
| 糖タンパク質のERADにおける、EDEMホモログタンパク質とER ManIの機能 EDEM1,2,3は、いずれもミスフォールドした糖タンパク質のERADを促進する働きをもっている(1,2,3)。また、ER ManIも哺乳類培養細胞にトランスフェクトして過剰発現させると、ミスフォールドした糖タンパク質の分解を促進する。ER ManIは小胞体に存在するマンノーストリミング酵素であるが、糖タンパク質のERADに焦点を絞ってその機能をみると、この酵素によってN結合型糖鎖がMan9からMan8Bにトリミングされることによって、糖タンパク質の品質管理機構は、積極的なフォールディングから分解の経路へと切りかわると考えられる(QS-A00, QS-A01参照)。EDEMがレクチンとして働いてこの過程を促進するという模式図は、今まで考えられてきたN結合型糖鎖によるERQCモデルに具体的な分子を加えることによって、このモデルをより確実なものにしたように見える。しかしながら、最近、分子レベルでの作用機序についての研究が進むに従って、新たなquestionが生まれてきている。 EDEM3の機能解析とEDEMホモログタンパク質の位置づけ 私たちが最近クローニングしたEDEM3は、培養細胞にトランスフェクトすると、ミスフォールドした糖タンパク質のERADを促進するとともに、N結合型糖鎖のマンノーストリミングを促進するという機能をもっていた。EDEM3の、クラスI α-1,2マンノシダーゼに保存された、酵素活性に重要な酸性アミノ酸に変異を導入すると、基質のマンノーストリミングが進まなくなることから、EDEM3はin vivoで酵素として働いている可能性が示された(3)。現在私たちは、EDEM3のin vitroでの酵素活性の検出に取り組んでいる。詳細は省くが、酵母のEDEMホモログ(Htm1p/Mnl1p)や、哺乳類のEDEM1,2に関してはin vivo, in vitroでの酵素活性は検出されておらず、従ってこれらのタンパク質はレクチンとして働いているのではないかと考えられている。 では、なぜ、EDEMグループとして分類されるタンパク質の中に、マンノースをトリミングする活性を持つものと持たないものがあって、一方で明らかな酵素活性を示すER ManIとは別のグループを形成しているのだろう?このことに関して私たちの考えているひとつの仮説は、EDEMのα-マンノシダーゼ (glycosylhydrolase family 47)ドメインは、酵素とレクチンのふたつの側面をもちうるのではないか、というものである。似た配列であっても、ある配列は酵素活性をもち、少し異なった配列は酵素活性をもたない可能性が考えられる。あるいは、酵素活性が非常に弱いとそれはin vivoではレクチンとして働く可能性もあるのではないか。最近Parodiらによって次のような興味深い実験データと仮説が提出された(4)。EDEMの機能を考える上で非常に参考になるので簡単に紹介したい。分裂酵母(Schizosaccharomyces pombe)にはER α-mannosidase(ER Man)が存在するにもかかわらず、今まで酵素活性は検出されていなかった。今回彼らは、分裂酵母のER Man破壊株を作製して実験したところ、糖タンパク質のERADが阻害されたことから、この分子は酵素としてではなくレクチンとして働いてERADを促進しているのではないか、というのである。彼らもその論文中でHtm1p/Mnl1p/EDEMについて言及しているが、私たちは今後、in vitroでの生化学的な解析を進めることによってこの点を明らかにし、EDEMホモログの機能解明とERADにおける分子基盤の解析を進めたいと考えている。 |
||||||||||||||||||||
| 細川暢子(京都大学・再生医科学研究所・細胞機能調節学) | ||||||||||||||||||||
|
||||||||||||||||||||
| 2006年6月5日 | ||||||||||||||||||||
|
|
||||||||||||||||||||
|
||||||||||||||||||||