 |
モデル生物C. elegansのグリコサミノグリカンには、コンドロイチン(Chn)とヘパラン硫酸(HS)が存在する。ヒアルロナンは存在しない(1)。以下では、両者について簡単に解説する。
コンドロイチン (Chn)
線虫のコンドロイチンは、ほとんど硫酸化されておらず、硫酸化されたChn(N-アセチルガラクトサミン残基のC4位のヒドロキシ基が硫酸化されている)の量はヘパラン硫酸(HS)程度である。Chnの硫酸化は酸化ストレス耐性に関わっているが、その他の機能は不明である。神経回路網の形成や神経再生などに関与している可能性もあり、今後の研究課題である(2)。
線虫のChn合成には、sqv-5とmig-22遺伝子によってコードされているコンドロイチン合成酵素とコンドロイチン重合化因子がはたらいている。sqv-5遺伝子またはmig-22遺伝子の機能をRNAiや遺伝子破壊によって阻害すると、初期胚の細胞分裂(細胞質分裂と細胞核分裂の両方)が異常となる(3)。図1には、コンドロイチンプロテオグリカン(CPG)のタンパク質部分(コアタンパク質)をコードする遺伝子cpg-1とcpg-2を同時に阻害した際に見られた細胞分裂異常を示した。細胞膜は緑の蛍光色素で生体染色してあり(写真下段)、上段は微分干渉顕微鏡写真である。CPG-1とCPG-2のコアタンパク質合成を阻害すると、細胞の分裂溝が入りこんで多細胞胚になった胚(図1a)で、時間とともに分裂溝が徐々に退縮し(図1b, c, d)、最終的には図1aで見られた分裂溝が全て消えて一つの多核の巨大細胞になってしまう(図1e)。マウスの初期発生でもChn合成が初期胚の分裂に必須であることを示す実験結果が得られており、Chnは哺乳類においても初期胚発生に不可欠な分子であると考えられる。線虫のChn合成は、さらに世界最小の器官といわれる線虫の陰門vulvaの形成にも不可欠である(1)。
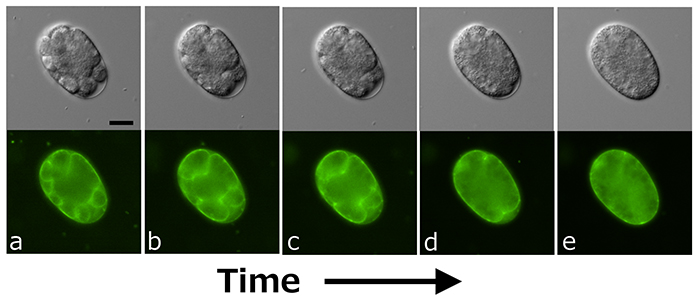
図 1. cpg-1遺伝子とcpg-2遺伝子を同時に阻害した際にみられる初期胚の細胞分裂異常
(詳細は本文参照。スケールバー 20 µm)
|
C. elegansでは現在、24種類のCPGのコアタンパク質遺伝子が同定されている(cpg-1からcpg-24)(4)。哺乳類と共通なドメインを有するものも数種類存在するが、細胞分裂に関与していることがわかっているのはcpg-1、cpg-2、cpg-14の3つの遺伝子である。CPG-1からCPG-24までの全タンパク質の立体構造(AlphaFold2 による予測)を図2に示す。ほとんどのコアタンパク質が天然変性領域intrinsically disordered region(IDR)とよばれる、一定の構造をとらない領域(図の緑の部分に含まれる)をもっており、Chnはこの部分に付加されていることが多い。CPG-1とCPG-2は、線虫の卵母細胞で表層顆粒とよばれるカベオリンを含む膜顆粒に存在しており、初期胚分裂の際には分裂溝に輸送される。両者のIDRをもつ立体構造(図3)は、液―液相分離で形成されるP顆粒と呼ばれる液滴(liquid droplet)構造を形成するタンパク質LAF-1と類似している(図3)。ヒトのプロテオグリカン(PG)であるシンデカン-4(SYND4)にもIDRがみられ(図3)、図2の多くの線虫のコアタンパク質(図3のCPG-9やCPG-19など)と類似している。SYND4は、HSとChnの両方の糖鎖修飾をうけるPGで、ヒトの培養細胞の細胞質分裂の進行に必須であり、midbody(中央体)に局在する(5)。IDRをもつコアタンパク質は、相分離でできる液滴の成分として、他の分子と相互作用する可能性がある。液滴に富む細胞核からの細胞分裂のシグナルが、CPGのコアタンパク質を含む液滴や膜で囲まれた表層顆粒の輸送とカップリングして細胞表面へと伝わっていく可能性もあり、今後の研究がまたれる。
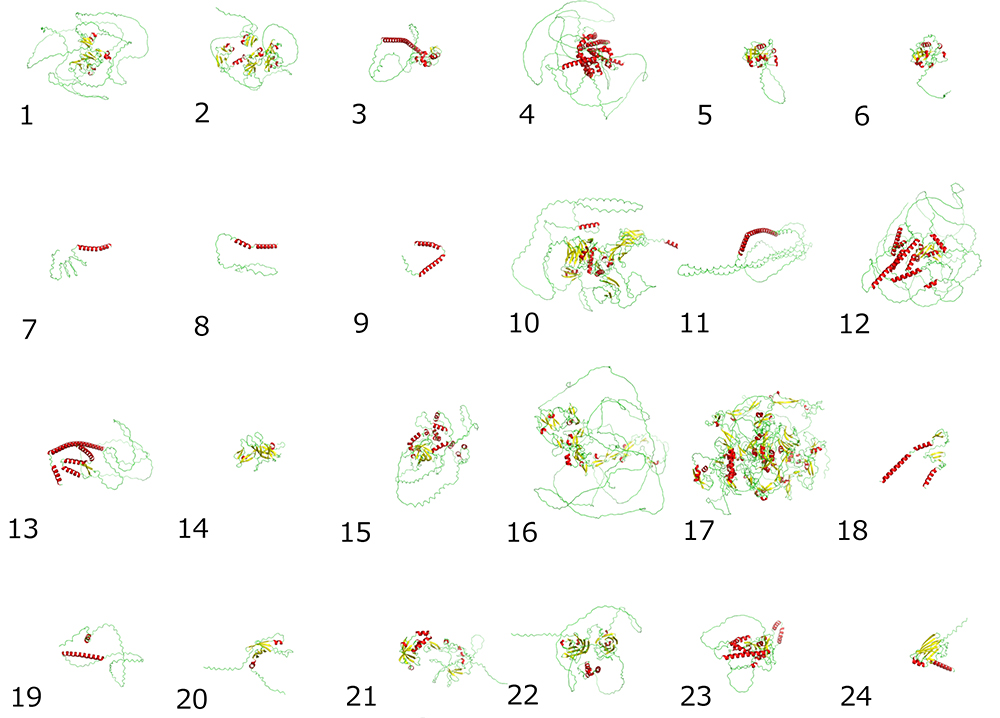
図 2. 24種のCPGの立体構造予測結果(AlphaFold Protein Structure Databaseより)
数字はcpg遺伝子の番号を示す。
|
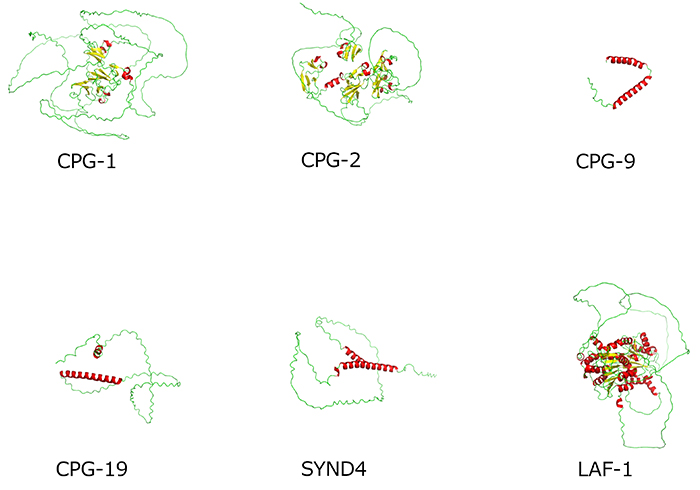
図 3. 天然変性領域IDRをもつと考えられているCPGタンパク質の立体構造の比較(AlphaFold Protein Structure Databaseより)
LAF-1はCPGではないことに注意。
|
ヘパラン硫酸 (HS)
線虫のHSはChnの0.2%程度の量を占めており、rib-1遺伝子(ヒトEXT1遺伝子のオーソログ)とrib-2遺伝子(ヒトEXTL3オーソログ)の遺伝子産物によって合成される。2つのタンパク質は複合体を形成してはたらくので、rib-1やrib-2遺伝子どちらかの機能を阻害するとHS合成が阻害される。発生後期の原腸陥入後の形態形成運動ventral enclosureの異常などで幼虫致死となったり、阻害が弱い場合には神経回路網形成の異常や咽頭形成の異常などが生ずる。これらの異常はそれぞれの段階ではたらくHSPGのコアタンパク質の合成阻害の効果と同じであり、またタンパク質全般の硫酸化を阻害した場合の異常と同じである。Chn合成阻害でみられた初期胚での細胞分裂異常は生じない(1)。HSPGのコアタンパク質としては、syndecan(sdn-1遺伝子産物)、glypicans(lon-2とgpn-1)、perlecan (unc-52)、agrin(agr-1)、type XVIII collagen(cle-1)が知られている。cle-1はcpg-10遺伝子でもある。
野村 一也(久留米大学医学部医化学講座)
| References |
| (1) |
Nomura K, Akiyoshi S, Matsuda A, Nomura KH: Glycosaminoglycans and glycosylphosphatidylinositol-anchor proteins in development of Caenorhabditis elegans. In Glycoscience: Biology and Medicine (Taniguchi N, Endo T, Hart GW, Seeberger PH, Wong, C-H eds), 817–824, Springer Japan, Tokyo, 2015 |
| (2) |
Izumikawa T, Dejima K, Watamoto Y, Nomura KH, Kanaki N, Rikitake M, Tou M, Murata D, Yanagita E, Kano A, Mitani S, Nomura K, Kitagawa H: Chondroitin 4-O-sulfotransferase is indispensable for sulfation of chondroitin and plays an important role in maintaining normal life span and oxidative stress responses in nematodes. J. Biol. Chem. 291, 23294–23304, 2016 |
| (3) |
Mizuguchi S, Uyama T, Kitagawa H, Nomura KH, Dejima K, Gengyo-Ando K, Mitani S, Sugahara K, Nomura K: Chondroitin proteoglycans are involved in cell division of Caenorhabditis elegans. Nature 423, 443–448, 2003 |
| (4) |
Noborn F, Larson G: Characterization of C. elegans chondroitin proteoglycans and their large functional and structural heterogeneity; evolutionary aspects on structural differences between humans and the nematode. Adv. Exp. Med. Biol. 21, 155–170, 2021 |
| (5) |
Addi C, Presle A, Frémont S, Cuvelier F, Rocancourt M, Milin F, Schmutz S, Chamot-Rooke J, Douché T, Duchateau M, Giai Gianetto Q, Salles A, Ménager H, Matondo M, Zimmermann P, Gupta-Rossi N, Echard A: The Flemmingsome reveals an ESCRT-to-membrane coupling via ALIX/syntenin/syndecan-4 required for completion of cytokinesis. Nat. Commun. 11, 1941, 2020 |
2023年 6月15日
|
|---|





