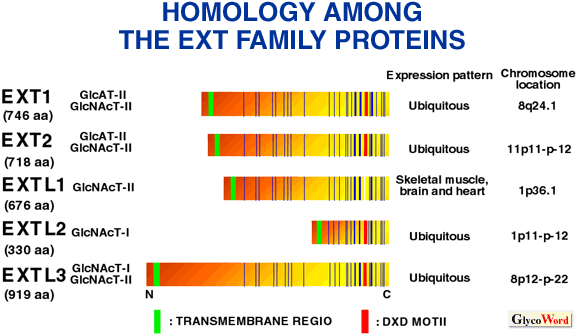
|
| References | (1) | McCormick C, Tufaro F : New perspectives on the molecular basis of hereditary bone tumours. Molecular Medicine Today 5, 481-486, 1999
|
|
(2) |
Sugahara K, Kitagawa H : Recent advances in the study of the biosynthesis and functions of sulfated glycosaminoglycans. Curr. Opin. Struct. Biol. 10, 518-527, 2000
|
|
(3) |
McCormick C, Leduc Y, Martindale D, Mattison K, Esford LE, Dyer AP, Tufaro F : The putative tumour suppressor EXT1 alters the expression of cell-surface heparan sulfate. Nat. Genet. 19, 158-161, 1998
|
|
(4) |
Lind T, Tufaro F, McCormick C, Lindahl U, Lidholt K : The putative tumor suppressors EXT1 and EXT2 are glycosyltransferases required for the biosynthesis of heparan sulfate. J. Biol. Chem. 273, 26265-26268, 1998
|
|
(5) |
Bellaiche Y, The I, Perrimon N : Tout-velu is a Drosophila homoloue of the putative tumor suppressor EXT-1 and is needed for Hh diffusion. Nature 394, 85-88, 1998
|
|
(6) |
Senay C, Lind T, Muguruma K, Tone Y, Kitagawa H, Sugahara K, Lidholt K, Lindahl U, Kusche-Gullberg M : Association of the EXT1 and EXT2 proteins in heparan sulfate biosynthesis. EMBO reports 1, 282-286, 2000
|
|
(7) |
Kitagawa H, Shimakawa H, Sugahara K : The tumor suppressor EXT-like gene EXTL2 encodes an alpha1,4-N-acetylhexosaminyltransferase that transfers N-acetylgalactosamine and N-acetylglucosamine to the common glycosaminoglycan-protein linkage region. The key enzyme for the chain initiation of heparan sulfate. J. Biol. Chem. 274, 13933-13937, 1999
|
|
(8) |
Kim B-T, Kitagawa H, Tamura J, Saito T, Kusche-Gullberg M, Lindahl U, Sugahara, K : The human tumor suppressor EXT gene family members, EXTL1 and EXTL3, encode alpha1,4-N-acetylglucosaminyltransferases involved in heparan sulfate/heparin biosynthesis. Proc. Natl. Acad. Sci. USA, in press, 2001
|
|
(9) |
McCormick C, Duncan G, Goutsos KT, Tufaro F : The putative tumor suppressors EXT1 and EXT2 form a stable complex that accumulates in the Golgi apparatus and catalyzes the synthesis of heparan sulfate. Proc. Natl. Acad. Sci. USA 97, 668-673, 2000
|
|
(10) |
Kobayashi S, Morimoto K, Shimizu T, Takahashi M, Kurosawa H, Shirasawa T : Association of EXT1 and EXT2, hereditary multiple exostoses gene products, in Golgi apparatus. Biochem. Biophys. Res. Commun. 268, 860-867, 2000
|
|
(11) |
Perrimon N, Bernfield M : Specificities of heparan sulfate proteoglycans in developmental processes. Nature 404, 725-728, 2000
|
|
(12) |
Lin X, Wei G, Shi Z, Dryer L, Esko JD, Wells DE, Matzuk MM : Disruption of gastrulation and heparan sulfate biosynthesis in EXT1-deficient mice. Dev. Biol. 22, 299-311, 2000
|
|
(13) |
Yamada Y, Van Die I, Van den Eijnden DH, Yokota A, Kitagawa H, Sugahara K : Demonstration of glycosaminoglycans in Caenorhabditis elegans. FEBS Letters 459, 327-331, 1999
|
|
(14) |
Toyoda H, Kinoshita-Toyoda A, Selleck SB : Structural analysis of glycosaminoglycans in Drosophila and Caenorhabditis elegans and demonstration that tout-velu, a Drosophila gene related to EXT tumor suppressors, affects heparan sulfate in vivo. J Biol Chem 275, 2269-2275, 2000
|
|
(15) |
Kitagawa H, Egusa N, Tamura J, Kusche-Gullberg M, Lindahl U, Sugahara K : rib-2, a Caenorhabditis elegans homolog of the human tumor suppressor EXT genes encodes a novel alpha1,4-N-acetylglucosaminyltransferase involved in the biosynthetic initiation and elongation of heparan sulfate. J. Biol. Chem. 276, 4834-4388, 2001
|
| |