|
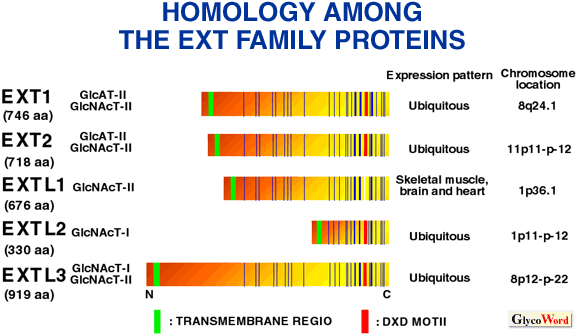
EXTL1, EXTL2 and EXTL3 also encode GlcNAc transferases, which are likely involved in HS biosynthesis (7,8). Truncated forms of EXTL1, EXTL2 and EXTL3, lacking the putative NH2-terminal transmembrane and cytoplasmic domains, were transiently expressed in COS-1 cells, and found to harbor alpha-GlcNAc transferase activities. EXTL2 protein transfers alpha1,4GlcNAc to GlcAbeta1-3Galbeta1-O-naphtalenmethanol (7), which is an artificial analog of the glycosaminoglycan-protein linkage region (GlcAbeta1-3Galbeta1-3Galbeta1-4Xylbeta1-O-Ser) and is a strong candidate for the key enzyme GlcNAcT-I that initiates the synthesis of HS and heparin (Hep), segregating it from the synthesis of chondroitin sulfate and dermatan sulfate, which are formed on the same linkage region structure. Truncated EXTL1 transfers alpha1,4GlcNAc to N-acetylheparosan oligosaccharides GlcAbeta1-4GlcNAcalpha1-(4GlcAbeta1-4GlcNAcalpha1-)n that represent growing HS chains, thus has GlcNAcT-II activity (8). Truncated EXTL3 utilizes not only N-acetylheparosan oligosaccharides but also GlcAbeta1-3Galbeta1-O-C2H4NHCbz, another synthetic substrate for alpha-GlcNAc transferase I for the initiation of the HS/Hep synthesis, thus has both GlcNAcT-I and GlcNAcT-II activities (8). Neither EXTL1 nor EXTL3 shows any glucuronyltransferase activity. Hence, EXTL3 is most likely involved in both chain initiation and elongation, whereas EXTL1 is possibly involved only in the chain elongation of HS and maybe Hep as well. Thus, the acceptor specificities of the five family members overlap, but are distinct from each other except for EXT1 and EXT2 with the same specificity. In other words, all five cloned human EXT gene family proteins harbor glycosyltransferase activities which probably contribute to the synthesis of HS and Hep.
A possible explanation for the existence of the two distinct GlcNAcT-I molecular species, EXTL2 and EXTL3, for the key enzyme activity critical for the selective assembly of HS/Hep chains may be that they initiate HS and/or Hep chains on different core proteins by discriminating the amino acid sequences. It remains to be clarified how the GlcNAcT-II activity of EXTL1 or the similar GlcNAcT-II activity of EXTL3 play their respective roles in chain polymerization of HS/Hep, where it requires a glucuronyltransferase (GlcAT-II) as a partner. It is unknown which one of EXT1 or EXT2 co-operates with EXTL1 and/or EXTL3 for the synthesis of the repeating disaccharide region or chain polymerization. Human EXT1 and EXT2 form a stable complex that accumulates in the Golgi apparatus (6,9,10) to exhibit substantially higher glycosyltransferase activities than EXT1 or EXT2 alone and appears to be a biologically relevant enzyme form (6,9). It should be remembered that HS chain polymerizing enzyme activity has not been demonstrated in vitro for an EXT1/EXT2 hetero-oligomeric complex. Therefore, possibility of multimeric complex formation involving other proteins also exists. Alternatively, the single GlcNAc transfer catalyzed by EXTL1 and EXTL3 possibly breaks up the elongation process and, in effect, serves as a chain termination mechanism.
The EXT and EXTL proteins are highly conserved from Caenorhabditis elegans and Drosophila melanogaster to higher vertebrates, and recent results from Drosophila genetics strongly indicate the essential roles of HS-synthesizing enzymes in developmental processes (11). Targeted disruption of EXT1 in mice results in embryonic lethality (12). HS has indeed been demonstrated in C. elegans (13,14) and Drosophila (14). Ttv is a protein of 760 amino acids and is 56% identical to the human EXT1 protein (5). The Hedgehog, but not FGF or Wingless, signaling, is selectively affected in the ttv mutant, suggesting that other EXT genes exist (5). Although ttv has been presumed to encode a putative HS polymerase, such catalytic activities of ttv or other Drosophila EXT proteins have not been reported. Two EXT genes designated rib-1 and rib-2 exist in C. elegans, and the rib-2 protein consisted of 814 amino acids has been shown to harbor GlcNAcT-I and GlcNAc-T-II activities, and rib-2 is an EXTL3 ortholog (15). No glucuronyltransferase activity involved in HS chain polymerization has been demonstrated for either protein.
|
|
|
| References | (1) | McCormick C, Tufaro F : New perspectives on the molecular basis of hereditary bone tumours. Molecular Medicine Today 5, 481-486, 1999
|
|
(2) |
Sugahara K, Kitagawa H : Recent advances in the study of the biosynthesis and functions of sulfated glycosaminoglycans. Curr. Opin. Struct. Biol. 10, 518-527, 2000
|
|
(3) |
McCormick C, Leduc Y, Martindale D, Mattison K, Esford LE, Dyer AP, Tufaro F : The putative tumour suppressor EXT1 alters the expression of cell-surface heparan sulfate. Nat. Genet. 19, 158-161, 1998
|
|
(4) |
Lind T, Tufaro F, McCormick C, Lindahl U, Lidholt K : The putative tumor suppressors EXT1 and EXT2 are glycosyltransferases required for the biosynthesis of heparan sulfate. J. Biol. Chem. 273, 26265-26268, 1998
|
|
(5) |
Bellaiche Y, The I, Perrimon N : Tout-velu is a Drosophila homoloue of the putative tumor suppressor EXT-1 and is needed for Hh diffusion. Nature 394, 85-88, 1998
|
|
(6) |
Senay C, Lind T, Muguruma K, Tone Y, Kitagawa H, Sugahara K, Lidholt K, Lindahl U, Kusche-Gullberg M : Association of the EXT1 and EXT2 proteins in heparan sulfate biosynthesis. EMBO reports 1, 282-286, 2000
|
|
(7) |
Kitagawa H, Shimakawa H, Sugahara K : The tumor suppressor EXT-like gene EXTL2 encodes an alpha1,4-N-acetylhexosaminyltransferase that transfers N-acetylgalactosamine and N-acetylglucosamine to the common glycosaminoglycan-protein linkage region. The key enzyme for the chain initiation of heparan sulfate. J. Biol. Chem. 274, 13933-13937, 1999
|
|
(8) |
Kim B-T, Kitagawa H, Tamura J, Saito T, Kusche-Gullberg M, Lindahl U, Sugahara, K : The human tumor suppressor EXT gene family members, EXTL1 and EXTL3, encode alpha1,4-N-acetylglucosaminyltransferases involved in heparan sulfate/heparin biosynthesis. Proc. Natl. Acad. Sci. USA, in press, 2001
|
|
(9) |
McCormick C, Duncan G, Goutsos KT, Tufaro F : The putative tumor suppressors EXT1 and EXT2 form a stable complex that accumulates in the Golgi apparatus and catalyzes the synthesis of heparan sulfate. Proc. Natl. Acad. Sci. USA 97, 668-673, 2000
|
|
(10) |
Kobayashi S, Morimoto K, Shimizu T, Takahashi M, Kurosawa H, Shirasawa T : Association of EXT1 and EXT2, hereditary multiple exostoses gene products, in Golgi apparatus. Biochem. Biophys. Res. Commun. 268, 860-867, 2000
|
|
(11) |
Perrimon N, Bernfield M : Specificities of heparan sulfate proteoglycans in developmental processes. Nature 404, 725-728, 2000
|
|
(12) |
Lin X, Wei G, Shi Z, Dryer L, Esko JD, Wells DE, Matzuk MM : Disruption of gastrulation and heparan sulfate biosynthesis in EXT1-deficient mice. Dev. Biol. 22, 299-311, 2000
|
|
(13) |
Yamada Y, Van Die I, Van den Eijnden DH, Yokota A, Kitagawa H, Sugahara K : Demonstration of glycosaminoglycans in Caenorhabditis elegans. FEBS Letters 459, 327-331, 1999
|
|
(14) |
Toyoda H, Kinoshita-Toyoda A, Selleck SB : Structural analysis of glycosaminoglycans in Drosophila and Caenorhabditis elegans and demonstration that tout-velu, a Drosophila gene related to EXT tumor suppressors, affects heparan sulfate in vivo. J Biol Chem 275, 2269-2275, 2000
|
|
(15) |
Kitagawa H, Egusa N, Tamura J, Kusche-Gullberg M, Lindahl U, Sugahara K : rib-2, a Caenorhabditis elegans homolog of the human tumor suppressor EXT genes encodes a novel alpha1,4-N-acetylglucosaminyltransferase involved in the biosynthetic initiation and elongation of heparan sulfate. J. Biol. Chem. 276, 4834-4388, 2001
|
| |