

|
|
Brain Development and Proteoglycans |
 |
||||||||||||||||
 |
The occurrence of multiple molecular species of proteoglycan is characteristic of the brain (Table 1). The detailed structure, expression pattern and functional role of an individual proteoglycan have recently been reviewed in several papers (1-4). This communication is a brief summary of the proposed functions of nervous tissue proteoglycans in the development of the brain. Proteoglycans, a group of glycoproteins that have covalently bound sulfated glycosaminoglycan chains, display affinity to various extracellular matrix constituents, cell adhesion molecules, or growth factors. Since each nervous tissue proteoglycan and its binding partners are usually co-localized in the brain, proteoglycans are considered to interact with these extracellular or cell surface molecules in the brain. Many cell culture studies have shown that neural proteoglycans are non-permissive molecules for axonal growth. For example, although neurite outgrowth from various neuronal cells is promoted when cells are cultured on dishes coated with laminin or L1/Ng-CAM, the co-existence of proteoglycans such as neurocan, phosphacan and agrin with laminin or L1 in the substrata results in neutralization of the promoting effects, even in inhibition of neurite outgrowth. These findings are consistent with the fact that proteoglycans exist in glial boundaries which prevent axonal elongation. In contrast, some studies revealed that neurocan or phosphacan promotes neuritogenesis of a certain class of neurons when cells are cultured on dishes coated with either of the proteoglycans. Additionally, in the developing cerebrum, the thalamocortical fibers grow in the subplate which contains a high concentration of neurocan, suggesting that neurocan can be a permissive molecule. Effects (inhibitory or promoting) of proteoglycans on the neurite outgrowth would depend on molecular species and/or composition of cell surface molecules which interact with the proteoglycans in the microenvironment of neurons. Proteoglycans also play an important role in growth factor signaling. Cell surface heparan sulfate proteoglycans including glypican and N-syndecan serve as a co-receptor for heparin-binding growth factors such as bFGF and pleiotrophin/HB-GAM, which are mitogenic and neurite-promoting factors in the brain. Evidence has accumulated to indicate that transmembrane chondroitin sulfate proteoglycans are involved in signal transduction as well. NG2 proteoglycan, expressed on glial progenitor cells, forms a complex with PDGF alpha-receptor and participates in PDGF signaling to promote the proliferation of these cells. RPTP zeta/beta is a brain-specific receptor-type tyrosine phosphatase bearing chondroitin sulfate chains, and could be involved in a certain signaling pathway. Neuroglycan C (NGC)/CALEB is a novel proteoglycan with both an acidic amino acid cluster and an EGF-like module. Since the amino acid sequence of its EGF-like module has a common feature of bioactive EGF molecules, it may be a new member of the EGF superfamily. One of the most attractive themes in neuroscience is the elucidation of the molecular mechanism governing neuronal activity-dependent maturation of brain function. In the early postnatal period, Cat-301 proteoglycan and neurocan are expressed in an activity-dependent manner in the visual and somatosensory systems, respectively. The activity-dependent expression of these proteoglycans could be a crucial step in the functional maturation of the brain. A number of studies have demonstrated that neural proteoglycans are involved not only in brain development as discussed above but also in neuronal regeneration in injured nervous tissues and in deposit-formation in the Alzheimer brain. Proteoglycans, which had been largely disregarded in neuroscience until the mid-1980s, are now coming into the limelight as bioactive nervous tissue substances. |
||||||||||||||||
|
|||||||||||||||||
| Atsuhiko Oohira (Institute for Developmental Research, Aichi) | |||||||||||||||||
|
|||||||||||||||||
| Mar.15, 1998 | |||||||||||||||||
|
|
|||||||||||||||||
|
|||||||||||||||||
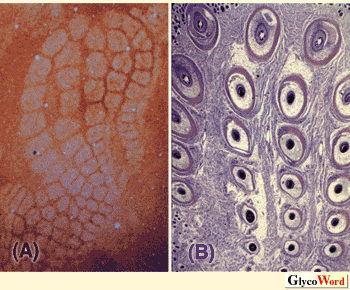 (A)Immunolocalization of neurocan to the septa of the barrels.
(A)Immunolocalization of neurocan to the septa of the barrels.
