|
|
グリコサミノグリカンとコアタンパク質との橋渡し領域の構造多様性 | |||||||||||||||||||||||
 |
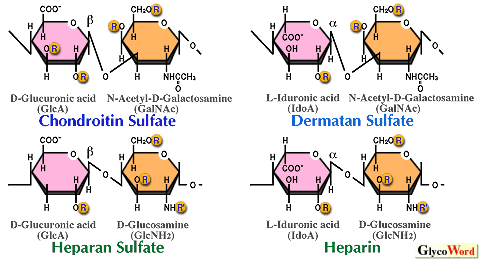
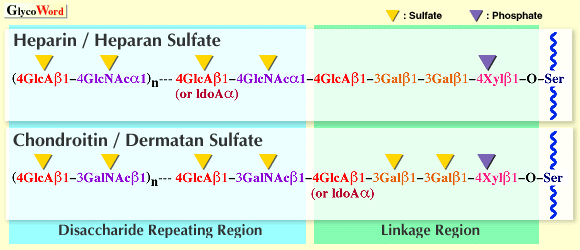
1)硫酸化グリコサミノグリカンの基本構造 プロテオグリカンの糖鎖部分である硫酸化グリコサミノグリカン(GAG)のうち、コンドロイチン硫酸、デルマタン硫酸、ヘパリン、ヘパラン硫酸は異なる構造と機能を有しているが、いずれも分子量数万の直鎖状の硫酸化多糖鎖である。糖鎖の大部分を占める領域はいずれのGAG鎖の場合もウロン酸(D-グルクロン酸またはL-イズロン酸)とアミノ糖(D-ガラクトサミンまたはD-グルコサミン)で構成されるニ糖単位が数十回繰り返した構造をしており、ニ糖繰り返し領域と呼ばれている(図1)。コンドロイチン硫酸とデルマタン硫酸はガラクトサミンを含むのでガラクトサミノグリカン、ヘパリンとヘパラン硫酸はグルコサミンを含むのでグルコサミノグリカンと呼べる。上記のような長いGAG鎖は繰り返しニ糖単位の構造が大きく異なるにも係わらず(図2)、いわゆる橋渡し領域といわれる共通の四糖(後述)を介して、コアタンパク質のGAG結合コンセンサス配列(Glu/Asp-X-Ser-Gly)中のセリン残基に共有結合している。 | ||||||||||||||||||||||
| |||||||||||||||||||||||
|
2)ニ糖繰り返し領域の多様性と機能ドメイン ニ糖繰り返し領域の構成単糖残基の硫酸化されうる部位は図2に示すように、ウロン酸側に1~2箇所、アミノ糖側に2~3箇所存在する。しかし、これらの部位は常に硫酸化されているわけではない。その結果、硫酸化パターンの違いによるニ糖単位の種類は理論的には16~48種類も存在しうることになる。機能ドメインは通常五~十糖の長さなので、仮に六糖構造を想定しても少なくとも16の3乗=4096種類の異なる硫酸化パターンが存在することになる。実際は、生合成の酵素の特異性によって限定されるので、天然に存在する構造の種類は幾分少なくはなるであろうが、いずれにしても極めて多種類の構造が存在し、その構造の多様性がGAG鎖の多様な生物学 的機能に対応していると考えられる。これらのGAG鎖は種々の機能性タンパク質と相互作用することによって、それらのタンパク質の機能発現の調節に関わっているのであろう。ちなみに、ヘパリンの血液凝固阻止活性はヘパリン上の特異的な硫酸化パターンを有したペンタサッカライド構造GlcN(6S)-GlcA-GlcN(NS,3S)-IdoA(2S)-GlcN(NS,6S) (NS, 2S, 3Sはアミノ基の硫酸化,2位,3位の水酸基の硫酸化)を介して、血中タンパク質であるアンチトロンビンIIIと特異的に結合することによって引き起こされる。塩基性繊維芽細胞増殖因子(basic FGF)と特異的結合活性を有するヘパラン硫酸上の五糖構造はGlcA-GlcN(NS)-HexA-GlcN(NS)-IdoA(2S) (HexA, GlcA or IdoA)であり、デルマタン硫酸の血液凝固阻止活性の基本となるヘパリンコファクターIIとの特異的結合ドメインはIdoA(2S)-GalNAc(4S)-IdoA(2S)-GalNAc(4S)-IdoA(2S)-GalNAc(4S)という六糖である。 3)タンパク質への橋渡し領域の共通四糖構造と修飾構造の多様性 コンドロイチン硫酸、デルマタン硫酸、ヘパリン、ヘパラン硫酸のニ糖繰り返し単位の構造は異なるが、面白いことにこれらのいずれのGAG鎖も共通の四糖橋渡し構造(GlcAβ1-3Galβ1-3Galβ1-4Xylβ1-)を介してコアタンパク質上の特定のSer残基の水酸基に共有結合している。この驚くべき事実は、1960年代に、酵素分解と部分酸加水分解の後に一群の中性糖ペプチドを単離して、それらの構造の比較から決定された(1)。1984年、ラットコンドロサルコーマのコンドロイチン硫酸プロテオグリカンの32Pによる代謝標識実験から、コンドロイチン硫酸鎖の橋渡し領域のキシロース残基の2位の水酸基がリン酸化されていることが示された(2)。翌年、ウシ肺由来のヘパラン硫酸に、その後サメ軟骨コンドロイチン硫酸などの橋渡し領域にも同じ構造が見い出された(表1)。1988年、ラットコンドロサルコーマ由来のコンドロイチン硫酸プロテオグリカンの橋渡し領域から単離されたグリコセリンにキシロースから数えて3番目のガラクトース(Gal-3)の4位の硫酸化が見い出された(3)。その後、同じガラクトース4-硫酸構造はクジラ軟骨など数種の異なる非ガン組織由来のコンドロイチン硫酸やウシ動脈由来のデルマタン硫酸の橋渡し領域にも見い出された(表1)。さらに、サメ軟骨のコンドロイチン硫酸に関する系統的な構造解析の結果、キシロースに隣接したガラクトース残基(Gal-2)の6位の硫酸化やGal-3の6位の硫酸化も見い出され、これらの硫酸化構造と上記のガラクトース4-硫酸構造の間の多くの組み合わせのパターンが存在することが判明した(4)。しかし、面白いことに、インターα-トリプシンインヒビターや尿中トリプシンインヒビター(一本のコンドロイチン硫酸鎖しか含まない)の場合には、必ずGal-3の4位の硫酸化がみられる。また、少なくともこれまでには、ヘパリンとヘパラン硫酸の橋渡し領域にはどのガラクトースの硫酸化構造も見い出されていない(表2)。キシロースのリン酸化とガラクトースの硫酸化が同じ糖鎖上に同時に見い出されたこともない。 4)橋渡し領域の修飾構造の生物学的意義は未解明の問題である 橋渡し領域の修飾構造の意義は不明であるが、いくつかの可能性が指摘されている。キシロースのリン酸化はERかゴルジ装置で起きているようであるが、培養繊維芽細胞ではリン酸化はGal-Gal-Xylが合成される間に起こり、次のGlcAの転移とともに急速な脱リン酸化が起こると報告されている。従って、プロテオグリカンの細胞内輸送に係わるシグナルであるか、生合成のなんらかのシグナルである可能性もある。ガラクトースの硫酸化が細胞内のどのオルガネラで行われているかはまだ調べられていない。また、リン酸基転移酵素や硫酸基転移酵素も同定されていない。しかし、生合成のシグナルである可能性が指摘されている(3,4)。橋渡し領域がまず構築されるので、その構造の違いはそれに引き続いて合成されるニ糖繰り返し領域の構造に影響することも推察される。GAG鎖の生合成のメカニズムに関する未解決の大きな命題は、ガラクトサミノグリカンとグルコサミノグリカンの仕分け合成の分子メカニズムの解明である。同じ橋渡し構造の上に合成されながら、機能も構造も異なるGAG鎖が合成される機構は依然として謎である。コアタンパク質上のアミノ酸配列の重要性も考えられているが(PG-A05参照)、橋渡し領域のガラクトースの硫酸化はガラクトサミノグリカンにしか見い出されていないので、橋渡し領域の四糖の次に転移されるGalNAcの未発見の転移酵素の認識部位になっている可能性も考えられる。キシロースのリン酸化のようにダイナミックな代謝回転をしている可能性も否定はできない。今後の研究が待たれる。 | |||||||||||||||||||||||
| 菅原一幸(神戸薬科大学・生化学教室) | |||||||||||||||||||||||
| |||||||||||||||||||||||
| 1998年 9月 15日 | |||||||||||||||||||||||
| |||||||||||||||||||||||