|
|
Structural Diversity of the Glycosaminoglycan-Protein Linkage Region | |||||||||||||||||||||||
 |
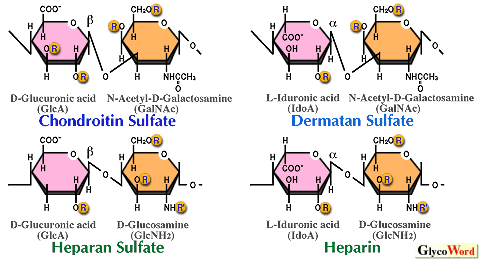
1) Structure of sulfated glycosaminoglycans (GAGs) Chondroitin sulfate (CS), dermatan sulfate (DS), heparin (HP) and heparan sulfate (HS), which are carbohydrate moieties of proteoglycans, have distinct structures and functions. They are linear sulfated polysaccharides with molecular weights of roughly 10,000 ~ 100,000. The large part of each GAG chain is constructed with a number of disaccharide repeating units and is called a disaccharide repeating region (Fig. 1). The repeating units are composed of uronic acid (GlcA or IdoA) and amino sugar (galactosamine or glucosamine). CS and DS that contain galactosamine are called galactosaminoglycans, whereas HP and HS that contain glucosamine are called glucosaminoglycans. Although all these GAG chain species have different repeating disaccharide regions, they are covalently bound through the so-called linkage region tetrasaccharide sequence (see below) to the serine residue in the GAG attachment consensus sequence (Glu/Asp-X-Ser-Gly) of respective core proteins. | ||||||||||||||||||||||
| |||||||||||||||||||||||
|
2) Structural diversity of repeating disaccharide regions and functional domains embedded therein One to 2 or 2 to 3 possible sulfation positions are found on the uronic acid and the amino sugar, respectively, in the repeating unit of each GAG species, as illustrated in Fig. 2. These positions are not always sulfated. Consequently, 16 ~ 48 different disaccharide units may exist depending on the combination of sulfation positions. Since functional domains are assumed to be usually penta- to decasaccharide long, at least 163 = 4096 different sulfation patterns on a hexasaccharide backbone for example are theoretically possible. Since, however, biosynthetic enzymes have restricted substrate specificities, the actual number of synthesized sequences in nature would be smaller. Even so, a large number of sequences still exist, and such a structural diversity corresponds to diversified biological functions of GAGs. These GAG chains are assumed to be often involved in modulating functions of various proteins such as growth factors and cytokines through specific interactions. For example, the anticoagulant activity of HP is exerted by the specific interaction of the HP pentasaccharide sequence GlcN(6S)-GlcA-GlcN(NS,3S)-IdoA(2S)-GlcN(NS,6S) (NS, 2S and 3S represent 2-N-, 2-O- and 3-O- sulfate, respectively) with antithrombin-III in blood. The pentasaccharide structure of HS responsible for the specific interaction with basic FGF has been proposed as GlcA-GlcN(NS)-HexA-GlcN(NS)-IdoA(2S) (HexA, GlcA or IdoA). The hexasaccharide sequence in DS that specifically binds to HP-cofactor II has been postulated as IdoA(2S)-GalNAc(4S)-IdoA(2S)-GalNAc(4S)-IdoA(2S)-GalNAc(4S). | |||||||||||||||||||||||
| |||||||||||||||||||||||
|
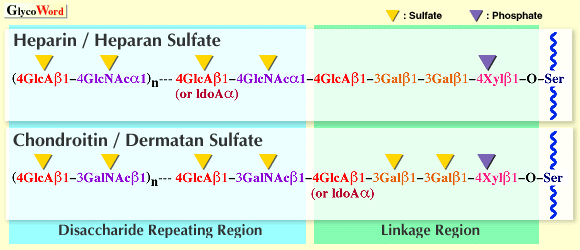
3) The common tetrasaccharide sequence in the GAG-protein linkage region and its multiple modifications Although disaccharide units of CS, DS, HP and HS are structurally different, every GAG chain of these is covalently bound to the OH group of the particular serine residue of core proteins through the linkage region tetrasaccharide sequence (GlcA beta1-3Galbeta1-3Galbeta1-4Xylbeta1-), which is common to different GAG species. This surprising finding was made in 1960s by structural comparison of the neutral glycopeptides isolated after enzymatic digestion and partial acid hydrolysis.(1) In 1984, C2 phosphorylation of Xyl in the CS linkage region was discovered by metabolic 32P-labeling experiments of rat chondrosarcoma proteoglycans.(2) An identical structure was found in bovine lung HS in the following year, and later in shark cartilage CS and so on (Table 1). In 1988, C4 sulfation of the Gal residue (Gal-3), which is in the third position from the reducing terminal Xyl, was discovered in a glycoserine isolated from the linkage region of rat chondrosarcoma CS.(3) An identical Gal-4-sulfate structure was found in galactosaminolgycan chains isolated from non-tumorous tissues as well such as whale cartilage CS and bovine aorta DS (Table 1). Furthermore, the systematic structural investigation of shark cartilage CS revealed C6 sulfation of Gal-3 as well as Gal-2 adjacent to the Xyl residue. Combinations of these and the above described C4 sulfation of Gal-3 displayed a number of different modification patterns in the linkage region.(4) Interestingly, however, the linkage region isolated from inter-alpha-trypsin inhibitor and urinary trypsin inhibitor, both of which bear a single CS side chain, was always sulfated on C4 of Gal-3. In contrast, neither Gal residue has been found to be sulfated in the linkage region of HP or HS (Table 2) (6) Phosphorylation of Xyl and sulfation of Gal have never been found concomitantly on a single GAG chain. 4) The biological significance of the modifications of the linkage region remains to be clarified. Although the significance of the modification of the linkage region has not yet been clarified, a few possibilities have been suggested. Phosphorylation of Xyl appears to take place in endoplasmic reticulum or Golgi's apparatus. In cultured fibroblasts, it has been demonstrated to occur while the linkage trisaccharide structure is being synthesized and addition of the first GlcA to the trisaccharide core is accompanied by rapid dephosphorylation. Hence, the transient 2-O-phosphorylation of Xyl may be a signal involved in intracellular transport and/or in the control of GAG biosynthesis. It has not been clarified in which intracellular organelle sulfation of the Gal residues takes place. The phosphotransferase or the sulfotransferases have not yet been identified either. However, it has been suggested that the sulfate groups on the Gal residues may be biosynthetic signals.(3,4) Since the linkage region is first constructed in biosynthesis, differences in the structure of this region may influence the structure of the repeating disaccharide region to be synthesized thereafter. One of the major unsolved problems in GAG biosynthesis is the molecular mechanism for biosynthetic sorting of galactosaminoglycans and glucosaminoglycans. It remains an enigma how GAG species that are different in their structures and functions can be synthesized on the common linkage region. The importance of the amino acid sequence in the core protein has been suggested (see PG-A05). On the other hand, since the sulfate groups on the Gal residues have been found only in galactosaminoglycans, they may well be recognized by the unidentified GalNAc transferase that adds the first GalNAc residue to the tetrasaccharide core sequence and leads to the synthesis of CS/DS. The possibility exists that sulfation and desulfation of the Gal residues may also occur in a dynamic fashion as in the case of phosphorylation and dephosphorylation of Xyl. Clarification of the biological roles of these phosphate and sulfate groups has to awaits further investigation. | |||||||||||||||||||||||
| Kazuyuki Sugahara (Department of Biochemistry, Kobe Pharmaceutical University) | |||||||||||||||||||||||
| |||||||||||||||||||||||
| Sep.15, 1998 | |||||||||||||||||||||||
| |||||||||||||||||||||||