

|
|
脳神経系におけるポリ(ジ、オリゴ)シアル酸 |
||||||||||||||||||||||||||
 |
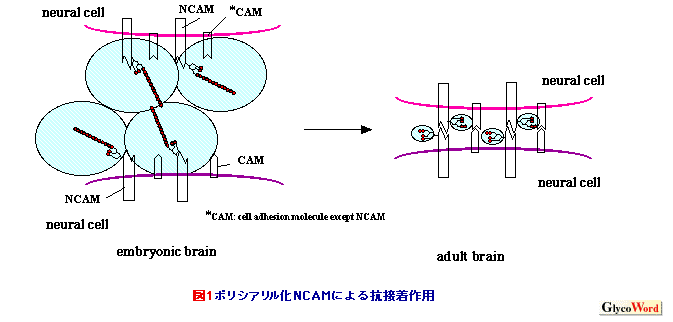
ポリシアル酸はシアル酸が重合度8-200程度で縮重合したポリアニオンである(1)。最もよく知られている構造はN-アセチルノイラミン酸(Neu5Ac)がα2,8結合で縮重合した重合体で、立体構造はN-アセチル基を外側にむけたヘリックス構造をとる。 |
|||||||||||||||||||||||||
|
||||||||||||||||||||||||||
|
|
||||||||||||||||||||||||||
| 近年、重合度がポリシアル酸よりも短い、ジ・オリゴシアル酸(重合度2-7)構造が様々な糖タンパク質上に証明され、ポリシアル酸との機能的差異が注目されている (3)。特にジシアル酸はb-シリーズガングリオシドのジシアル酸構造と同じ配列であり、ジシアリル化ガングリオシドとジシアリル化糖タンパク質の両方が共局在する脳、神経系においては、その機能の重要性が示唆される。神経分化を誘導できる培養細胞である神経芽腫細胞Neuro2Aを用いた解析から、ジシアル酸含有糖タンパク質として、免疫グロブリンスーパーファミリーの一員である接着分子CD166が同定され、ジシアル酸含有糖タンパク質のジシアル酸構造が神経突起形成に関わることが示された(4)。 | ||||||||||||||||||||||||||
| 佐藤ちひろ(名古屋大学大学院生命農学研究科) | ||||||||||||||||||||||||||
|
||||||||||||||||||||||||||
| 2004年11月30日 | ||||||||||||||||||||||||||
|
|
||||||||||||||||||||||||||
|
||||||||||||||||||||||||||

