

|
|
神経回路形成におけるコンドロイチン硫酸の役割 |
||||||||||||||||||||||||||
 |
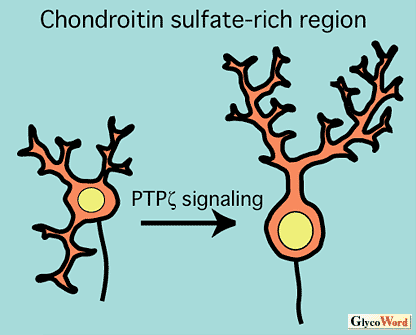
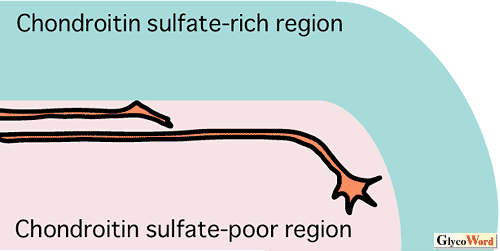
神経回路の形成過程において、各神経細胞は、特徴的な形態を発達させると共に、その軸索を特異的な経路に沿って伸長させることによって、正しい標的細胞を見出し、シナプスを形成する。これまで多くの研究者が、発達期脳組織をコンドロイチナーゼABCで処理し、コンドロイチン硫酸を分解除去すると、神経細胞の形態形成や軸索走行に大きな異常が生じることを見出している。このような観察から、コンドロイチン硫酸は、軸索伸長に対する反発性因子として軸索誘導に関与する(図B)、あるいは、神経細胞の形態分化に寄与するものと(図A)考えられている。最近、コンドロイチン硫酸の糖鎖構造依存的な情報伝達制御が、神経回路形成に重要な役割を果たしていることが明らかにされつつある。 |
|||||||||||||||||||||||||
|
||||||||||||||||||||||||||
|
||||||||||||||||||||||||||
 |
||||||||||||||||||||||||||
|
||||||||||||||||||||||||||
| PTPζは、コンドロイチン硫酸プロテオグリカンとして生合成される脳特異的な受容体型チロシンホスファターゼであり、その細胞外領域は脳の主要な可溶性コンドロイチン硫酸プロテオグリカンであるホスファカンとして分泌される。PTPζは、ヘパリン結合性成長因子であるプレイオトロフィンをリガンドとするが、両者の高親和性結合および情報伝達には、本受容体に結合しているD単位[GlcUA(2S)-GalNAc(6S)]を含むコンドロイチン硫酸の寄与が必要であると考えられる(1)。最近、筆者らは、PTPζとプレイオトロフィンの情報伝達が小脳プルキンエ細胞の形態形成に関与すること明らかにした(2)。すなわち、ラット小脳スライス培養系を用いた実験により、本情報伝達系を阻害すると、過剰な一次樹状突起や異常な方向に伸びる一次樹状突起をもつプルキンエ細胞が出現することを見出した(図A)。プルキンエ細胞の周囲には、D単位に富むコンドロイチン硫酸が豊富に存在し、コンドロイチン硫酸の糖鎖構造依存的な情報伝達の場として注目される。 一方、菅原らは、マウス16日目胚より調製した海馬神経細胞を、各種コンドロイチン硫酸(デルマタン硫酸)標品を塗布した基質上で培養することにより、特定の構造を有する糖鎖が、神経突起伸長促進作用を示すことを見出した(3)。中でも、DあるいはiD単位[GlcUA/IdoUA(2S)-GalNAc(6S)]を多く含むものは樹状突起様の突起を伸展させ、EあるいはiE単位[GlcUA/IdoUA-GalNAc(4S,6S)]に富むものは、長い軸索様の突起を伸長させる。このような高硫酸化されたコンドロイチン硫酸やデルマタン硫酸は、多くのヘパリン結合性成長因子と結合することが知られており、これらの情報伝達が海馬神経細胞の形態形成に関与していることが示唆される。 以上の例では、コンドロイチン硫酸は神経細胞に対して均一な環境因子として提示されているが、発達過程の脳においては、コンドロイチン硫酸がある種の境界面に発現することも多い。このようなコンドロイチン硫酸が豊富に存在する領域は軸索伸長に対して阻害的に作用し、軸索が誤った方向に侵入するのを妨げていると考えられている(図B)。マウス視交叉やニワトリ終脳−間脳境界における軸索ガイダンスがその例であり、このような領域をコンドロイチナーゼABCで処理すると、網膜神経節細胞の軸索伸長経路が大きく乱れることが知られている(4)。最近、境界面におけるコンドロイチン硫酸の濃度差のみならず、構造的な差異も軸索ガイダンスに寄与することが明らかにされつつある(4)。コンドロイチン硫酸は、軸索再生の阻害因子として、近年、注目を集めており、このような反発作用の分子機構の解明が大いに待たれるところである。 |
||||||||||||||||||||||||||
| 前田信明 (東京都神経科学総合研究所・分子発生生物学研究部門) | ||||||||||||||||||||||||||
|
||||||||||||||||||||||||||
| 2004年2月15日 | ||||||||||||||||||||||||||
|
|
||||||||||||||||||||||||||
|
||||||||||||||||||||||||||

