

 この作品はクリエイティブ・コモンズ 表示 4.0 |
 |
|---|
糖転移酵素の複合体形成による制御 | ||||||||||||||||||||||
 |
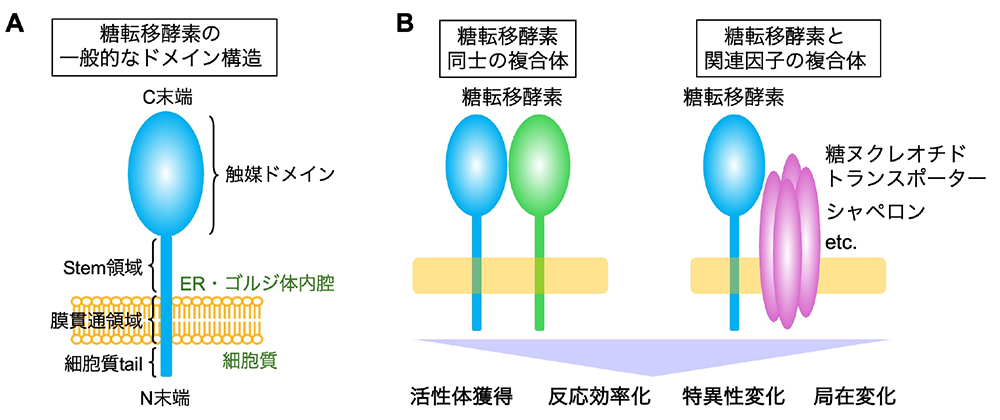
糖転移酵素は、糖鎖の生合成を触媒する酵素群であり、動物では約200種類が知られ、そのほとんどは小胞体(ER)、ゴルジ体に局在する膜タンパク質である(O-GlcNAc転移酵素OGTなどは例外的に他の場所に局在)。糖鎖の前駆体・根元やGlycosylphosphatidylinositol (GPI)などの生合成に関わる酵素は複数回膜貫通タンパク質であることが多いが、多くの糖転移酵素はII型の1回膜貫通型タンパク質であり、短いN末端領域をサイトゾルへ、触媒ドメインを含む大きなC末端領域をオルガネラ内腔側へ配向させたトポロジーをとる(図 A)。そしてこれらの酵素が段階的に作用することによって、多種多様な糖鎖構造が作られる。 ゴルジ体は、大別するとN型糖鎖、O型糖鎖、グリコサミノグリカン(GAG)、GPI、糖脂質、という5つのクラスの糖鎖を常時作り、非常に多様な構造の糖鎖を作り上げている。各クラス固有の糖転移酵素と、クラス間で共通の構造(末端が多い)を作る酵素が存在し、これらの酵素はゴルジ体の中で、他の酵素と時に競合、時に協調しながらその細胞固有の糖鎖パターンを作り上げる。この複雑な糖鎖生合成プロセスは、ただ酵素と基質がランダムに出会った結果起こるわけではなく、様々な因子により高度に制御されている。例えば、酵素の精密なゴルジ体内局在(糖転移酵素の局在の頁(近日公開予定)参照)や、酵素による特定のタンパク質基質の認識(“糖転移酵素の基質タンパク質選択性”の頁参照)の仕組みは、糖鎖の生合成に大きな影響を与える。そして、そのような制御因子の一つとして、酵素の複合体形成がある。 糖転移酵素は、様々なタンパク質と複合体を形成することが報告されている(ここでは同一酵素のホモオリゴマーを除く)。よく知られた例では、ある特定の糖鎖を合成する糖転移酵素同士が複合体を形成することで(図 B左)、その糖鎖を合成するために有利な活性や特異性を獲得するというものである。一部の例を紹介すると、例えば、ポリラクトサミン構造を作るB3GNT2とB3GNT8はそれぞれ単体でも活性を持つが、内腔側ドメインで複合体を形成することで約10倍、100倍と活性が上昇する(1)。またO-Man型糖鎖の最初のステップを担うPOMT1とPOMT2や、ヘパラン硫酸のポリマー化を担うEXT1とEXT2などは、両者が複合体を形成して初めて強い活性を発揮するようになる(2,3)。O-Man型糖鎖の生合成においては、POMGNT1とFKTNという、同一経路上の異なるステップを触媒する酵素同士の膜貫通ドメインを介した相互作用も報告されており(4)、同様にhuman natural killer-1 (HNK-1)糖鎖を段階的に作るのに必要なB4GALT2, GlcAT-P, HNK-1STが効率的合成を行うための複合体を形成することなども報告されている(5)。このように、一連の糖鎖合成経路上の酵素が複合体を形成する例が多く知られている。 一方、糖鎖合成酵素同士だけでなく、糖転移酵素と様々な関連因子とのヘテロ複合体も報告されている(図 B右)。例えば、B4GALT1がα-lactalbuminと結合することによって、基質特異性を変化させてラクトース合成酵素として機能することは古くからよく知られている(6)。またムチン型糖鎖合成に関わるC1GALT1は、特異的シャペロンであるCOSMCとの相互作用が活性体となるために不可欠である(7)。また、糖鎖の原料となる糖ヌクレオチドのトランスポーターと、その糖ヌクレオチドをドナー基質とする特定の糖転移酵素との複合体形成も知られている。古い例では、UDP-Galトランスポーターがガラクトシルセラミド(GalCer)合成酵素UGT8Aと複合体を形成してトランスポーターの局在を調節したり(8)、最近ではUDP-GlcNAcトランスポーターが特定のN型糖鎖分岐酵素と複合体を形成することも報告されている(9)。
一方で、こうした複合体形成については、細胞株を用いた過剰発現の系で証拠が得られている場合が多いため、内在性の酵素が組織中でどのように複合体を形成しているのかはいまだ十分な証拠が得られていないケースもある。近年、CRISPRを用いたゲノム編集、またTurboIDなどを始めとする近接標識などの技術が進展しており、内在性酵素へのタグのノックインや、IP(免疫沈降)-ウエスタンブロッティングに依らない相互作用の検出などが可能になってきている。これらの技術の進展とともに、様々な細胞において内在性の酵素がどのような複合体状態で存在しているのかを明らかにすることが、糖鎖生合成の全貌を理解する上で必要な次のステップになると考えられる。 木塚 康彦(岐阜大学 糖鎖生命コア研究所)
|
|||||||||||||||||||||
2024年 3月15日

