

|
|
ガレクチンの比較生化学 |
|||||||||||||||||
 |
ガレクチンはβ-ガラクトシドを認識する動物レクチンであり、これまでに線虫、ハエ、マアナゴ、ゼブラフィッシュ、ニジマス、カエル、ニワトリ等々、多様なモデル生物から同定され解析が進められている(「レクチン」のジャンル「LE-A01ガレクチン:定義と命名の経緯」参照)。哺乳類では現在14種が同定され、構造上三種のサブタイプに分類されているが(galectin-1〜14)、哺乳類以外の脊椎動物で包括的に同定、解析がなされているのは、我々がガレクチン研究のモデル生物として導入しているアフリカツメガエルのみである。そこで本稿では、アフリカツメガエルと哺乳類のガレクチンファミリーの比較について述べる。 | ||||||||||||||||
 |
|||||||||||||||||
|
|||||||||||||||||
|
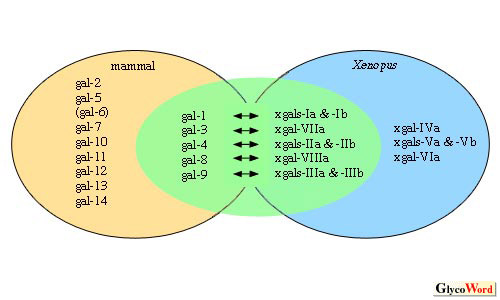
ツメガエル・ガレクチン(現在12種同定;xgalectin-Ia〜VIIIa)と哺乳類ガレクチンをアミノ酸配列および発現パターンの類似性の観点から比較してみると、図1に示す様に明らかな相同物とみなせるものと、みなせないものに分類される(1,2)。相同物と想定できるものについては、糖鎖認識活性の特異性も調べた一例では、類似していた(galectin-3とxgalectin-VIIa)。また、発現パターンが類似した代表例としては、galectin-4とxgalectins-IIa
& -IIb がともに消化管に局在することが挙げられる。しかしながら、それらの全長のアミノ酸配列の一致率は、いずれの組み合わせもほぼ50%前後にとどまっている。この値は、例えば生体の形成や生存に必須な転写因子群や増殖因子群について比較した場合に比べて、低いものである。こうした機能上普遍性が高いと考えられるタンパク質のアミノ酸配列は、カエルと哺乳類の間でも概ね7割程度以上は一致するものである。
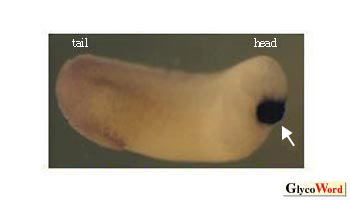
また、相同関係が推測できないものが多数存在するが、その一例として、哺乳類とツメガエルの皮膚特異的ガレクチンについて述べる。いずれの動物にも、皮膚に限局して多量に存在するガレクチンがある(哺乳類galectin-7、ツメガエルxgalectin-Va & Vb)。しかしそれらは、アミノ酸配列の比較からはおよそ相同的と呼べる類似性は見いだせない(一致率15%)。このことは、同じ皮膚でありながら、両動物種の皮膚は一見して異なっていることを反映している、あるいは各ガレクチンが、各動物種の皮膚を特徴づける一役割を担っているのではないかと想像させる。すなわち、相同関係が推測できるものでも、それらの全長アミノ酸配列の類似性はさほど高くないことも考慮に入れると、ガレクチンファミリーは、種を越えた普遍的な作用をもつ一方で、種を特徴づけることにも大いに関わっているのでは無いかと推測される。実際、様々な動物種のDNAデータベースが充実してきた今なお、galectin-7遺伝子は哺乳類にしか発見できない(3)。また、相同関係が見いだせないツメガエルxgalectin-VIa の、発生過程におけるmRNA の分布を追ったところ、両生類の胚にのみ一過的に形成されるセメント腺と呼ばれる組織に極めて特異的に発現していた(図2)。 |
|||||||||||||||||
 |
|||||||||||||||||
|
|||||||||||||||||
|
以上のことは、必ずしもガレクチンファミリーに限らず、糖鎖関連遺伝子全体の傾向として見てよいのではないかと筆者は考えている。糖鎖関連遺伝子、ひいては糖鎖そのものは、種を特徴づけることに大きく貢献し、さらには進化の原動力の一翼を担っているのではないだろうか。
|
|||||||||||||||||
| 東海林 博樹(香川大学医学部 分子細胞機能学) | |||||||||||||||||
|
|||||||||||||||||
| 2005年1月24日 | |||||||||||||||||
|
|
|||||||||||||||||
|
|||||||||||||||||

