|
|
植物 ADPglucose pyrophosphorylase の機能改変 |
 |
||||||||||||||||||||||||||||||||||||||||||||||||||||||
 |
ADPglucose は、細菌のグリコーゲン生合成や植物のデンプン生合成において、ATPと glucose-1-phosphate から合成されたのち、グルコース供与体として用いられる。この反応を触媒するのが、ADPglucose pyrophosphorylase (EC 2.7.7.27; 以下AGPase) である(式1)。 細菌と植物の AGPase はともに同じ反応を触媒するにも関わらず、両者の構造や調節機構は異なる。まず、細菌の AGPase が glgC 遺伝子にコードされるタンパク質からなるホモ4量体であるのに対して、植物の AGPase は異なる遺伝子にコードされる大サブユニットと小サブユニットからなるヘテロ4量体である。また、これまでに明らかにされた細菌の AGPases とほとんどの植物 AGPase は、アロステリック調節を受けているが、その調節特性は若干異なり、細菌の酵素はfructose-1,6-bisphosphate により活性化され、AMP で阻害される。一方植物の場合は、3-phosphoglycerate (3-PGA) で活性化され、Pi で阻害される(表1)。 |
||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||
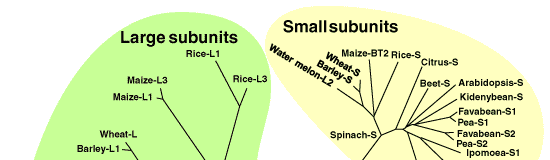
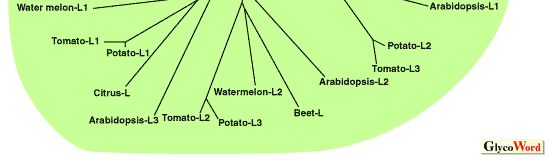
| 植物 AGPase の大小サブユニットをコードする多数の遺伝子配列がすでに明らかにされている。それらの一次構造から示唆されていることは、(1)小サブユニットは互いに非常に高い相同性(70%以上)を持つ、(2)大サブユニットも比較的高い相同性(50%以上)を有する、(3)大小サブユニット間にも相同性(40%以上)がある、ことである(図1)。一つの酵素における大小サブユニットの分子量はおおむね 50-60 kDa で、その差は小さいが、依然として大小を区別している。それぞれが異なる役割を有するがゆえである。アロステリック調節を受けるのが主として大サブユニットであるのに対して、酵素としての触媒活性を担うのが小サブユニットである [1]。このことは、小サブユニットの一次構造が高度に保存されていることからも理解できる。AGPase の構造と機能の関係は、主に大腸菌の酵素を材料に化学修飾法や部位特異的変異を用いて調べられ、基質結合領域やアロステリック分子の結合領域が同定されてきた。より複雑な構造の植物の AGPase に関する報告は非常に少ないが、それらの結合領域のアミノ酸配列は植物の酵素でも保存されていることから、同じく基質やアロステリック分子が結合すると推定される [1]。 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 植物における AGPase のアロステリック特性はデンプンの量や質を左右するという証拠が、植物変異体や形質転換植物を使ったデータから多数示されてきた。また、in vivo 部位特異的変異法によるトウモロコシ AGPase 大サブユニット遺伝子の変異は、Pi に対して非感受性の AGPase をもたらし、その結果種子重量が非形質転換体の 11-18% 増加することが示された [2]。このことは、AGPase の アロステリック変異酵素の植物への導入が 種子重量や収量を変えうる可能性を示している。したがって、植物 AGPase の立体構造は、現在のところ明らかにされていないが、酵素構造の理論的改変を導くに違いない。少々強引かもしれないが、ランダム変異法と部位特異的変異法を組み合わせて植物 AGPase の機能改変が試みられている。この方法により、アロステリック分子に非感受性のジャガイモ AGPase 変異酵素が作製された(表2)[3, 4]。たとえば、38 番目のグルタミンがリジンに変わった大サブユニットと野生型小サブユニットからなる変異 AGPase (UpReg-1) では、活性化に必要な3-PGA は野生型の 80分の1でよく、阻害に必要な Pi の濃度は野生型 AGPase の70倍になった。Arabidopsis AGPase 大サブユニット遺伝子の同じ位置に変異を導入した場合も、アロステリック分子に対して非感受性になった AGPase が形成されたことから、1アミノ酸置換で酵素構造が変化したものと考えられる。さらに、38番目のグルタミンをリジンに、101 番目のグリシンをアスパラギンに換えた二重変異体は、UpReg-1 や UpReg-2 以上にアロステリック非感受性になる可能性があり、現在解析を進めている。このような変異遺伝子を植物へ導入することによって、植物デンプン合成速度の変換がもたらされる可能性が多分にある。 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 伊藤浩之(北海道大学大学院農学研究科) | |||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 2000年 12月 15日 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||