|
|
β-アミラーゼの活性中心 |
 |
||||||||||
 |
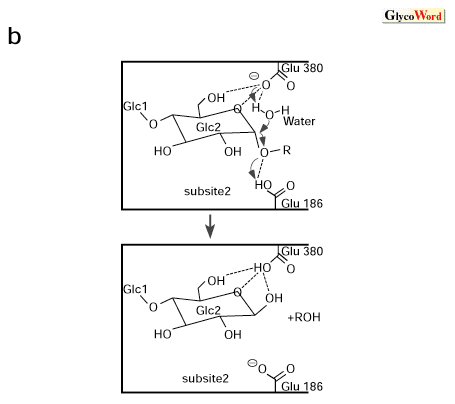
β-アミラーゼは澱粉の非還元末端からマルトース(麦芽糖)を遊離する酵素であり、生成物は常にβ-アノマーのマルトースである。酵素反応によってアノマー型の反転を伴い、Koshlandによってグルコアミラーゼとともに代表的な反転(inverting)酵素に分類されている。本酵素はダイズ、サツマイモ、オオムギなどの高等植物および一部の微生物に分布し、分子量は5-6万の単純タンパク質で構成されている。高等植物の酵素とバクテリア起源の酵素とは至適pHなどの性質に差があり、微生物起源の酵素が生澱粉を吸着、分解することができるのに対して、高等植物起源の酵素には生澱粉分解活性はない。 β-アミラーゼの立体構造はダイズ起源の酵素のX線結晶構造解析が最初に行われ、サツマイモ、オオムギおよびBacillus cereus起源の酵素の構造が明らかにされている。その結果、触媒ドメインは(β/α)8バレルから成り、β-バレルのC末端側に形成されるクレフトは(β/α)8バレルの3番目、4番目、5番目からのびる長いループによって取り囲まれ、深い活性部位のポケットを形成している。植物酵素のC末端はバレルを囲む長いループになっているが、Bacillus cereus起源の酵素ではこのループが澱粉吸着ドメインに置き換わっている、β-アミラーゼの(β/α)8バレル構造はα-アミラーゼファミリーの酵素の触媒ドメインを構成する(β/α)8バレルとは異なりヘリックスの数やループ構造に共通性は認められず、両者は異なるファミリーに属すると考えられている。 本酵素の反応機構はダイズ酵素とマルトースとの複合体のX線結晶構造解析の結果から、推定されている。マルトースは2分子が活性部位の触媒部位を挟んで結合するが、これは1分子のマルトースが生成物のβ-アノマーの形でサブサイト1-2に、もう1分子がサブサイト3-4に結合することで可能になっている。図1-aに2分子のマルトースと相互作用する酵素の側鎖を示した。サブサイト1から4までのサイトはグルコース残基と8-10本の水素結合を形成するのに対して、サブサイト4ではHis300との間に1本の水素結合しか形成せず、その代わりに周囲の疎水性残基と多くのファンデアワールス接触をしている。残基番号96から103の部分はフレキシブルループを形成し、シクロデキストリン複合型ではオープン構造をとるのに対して、マルトース複合体ではクローズド構造をとり、基質の結合に関与する。この複合体で注目すべき点はサブサイト2と3の間で糖鎖の2面角が180゜近く回転して結合していることであり、このことが触媒機構に深く関わっている。サブサイト2と3の間にある触媒部位にはGlu186の側鎖がサブサイト3のO1の近くに、Glu380の側鎖がサブサイト2のβ-アノマーのO1の近くに配置され、水素結合を形成している。 これらのことから本酵素の触媒機構は図1-bのように推定されている。Glu186は一般酸触媒として働き、グリコシド結合のOに水素を付加し、反対側からGlu380が一般塩基触媒として、触媒水から水素を引き抜き、活性化された水がサブサイト2のグルコース残基のC1原子を攻撃し、水酸基を付加する。この際、水分子の攻撃が常にβ-アノマーの位置からおこるため、生成物がβ-アノマーに固定されることになる。また、サブサイト2に結合しているグルコース残基の環はハーフチャアーに近い平面性を示すことから、反応の中間体として、カルボニウムカチオンの存在が推定される。この切断過程が終了し、フレキシブルループがオープン構造をとった時に生成物が放出され、次の基質が取り込まれると推定されている。 |
||||||||||
|
|||||||||||
|
|||||||||||
|
|||||||||||
| 三上文三 (京都大学食糧科学研究所) | |||||||||||
|
|||||||||||
| 2000年 6月 15日 | |||||||||||
|
|
|||||||||||
|
|||||||||||